Article Text

Abstract
Objective The aim of the study was to investigate whether the IRAK1/JAK2/Flt3 inhibitor pacritinib prevents disease development in the lupus-prone ABIN1[D485N] knock-in mouse.
Methods ABIN1[D485N] knock-in mice aged 8 weeks were fed for 10 weeks on a diet containing pacritinib. Body weight was monitored, and serum collected at the end to measure pacritinib, autoantibody and immunoglobulin levels. Splenic immune cell populations were analysed, and the kidney, liver and lungs examined for pathology.
Results Pacritinib prevented multiple facets of the lupus phenotype in ABIN1[D485N] knock-in mice, including splenomegaly, expansion of splenic germinal centre B cells, follicular T helper cells, and neutrophils, elevated serum levels of double-stranded DNA antibodies and immunoglobulins, glomerular IgA and lung inflammation.
Conclusions Pacritinib may be useful for the treatment of multiorgan inflammation in patients with lupus.
- Autoimmune Diseases
- Lupus Erythematosus, Systemic
- Lupus Nephritis
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
WHAT IS ALREADY KNOWN ON THIS TOPIC
The TLR7-MyD88-IRAK4-IRAK1 pathway has been shown to drive the lupus phenotype in the ubiquitin-binding deficient ABIN1[D485N] knock-in mice.
Crossing ABIN1[D485N] knock-in mice to interleukin-6 knock-out mice prevents the increase in serum levels of autoantibodies, immunoglobulin and kidney pathology, and crossing to mice expressing a kinase-inactive IRAK4 or an IRAK1 mutant suppresses all facets of the lupus phenotype.
WHAT THIS STUDY ADDS
We show that the JAK2/IRAK1/Flt3 inhibitor pacritinib, which has been approved for the treatment of myelofibrosis in humans, prevents lupus development in the ABIN1[D485N] knock-in mice.
HOW THIS STUDY MIGHT AFFECT RESEARCH, PRACTICE OR POLICY
The study suggests that pacritinib may have therapeutic potential for the treatment of lupus by targeting both the TLR7-MyD88-IRAK1 pathway and by limiting B cell numbers and function by inhibiting Flt3 and JAK2, respectively.
Introduction
Human polymorphisms in Tnip1 encoding the protein ABIN1 (A20-binding inhibitor of NF-κB 1) predispose to SLE in multiple human populations.1 2 ABIN1 exerts this effect by binding to particular types of ubiquitin oligomer, thereby restricting activation of the protein kinases TAK1 and IKKβ and the production of inflammatory mediators during toll-like receptor (TLR) signalling. To mimic the human condition, we generated knock-in mice expressing the ubiquitin-binding defective ABIN1[D485N] mutant instead of wild type (WT) ABIN1. These mice develop a lupus-like phenotype starting at 3 months of age, which is characterised by the presence of autoantibodies against double-stranded DNA (dsDNA) and anti-nuclear antigens, splenomegaly, increases in immune cell populations including monocytes, germinal centre B (GCB) cells, follicular T helper (TFH) cells and neutrophils, deposition of immunoglobulins in the kidneys and multiorgan inflammation.3 These phenotypes are abolished by crossing the ABIN1[D485N] mice to TLR7 knock-out (KO) mice, or myeloid differentiation primary response 88 (MyD88) KO mice or knock-in mice in which the protein kinases IRAK4 or IRAK1 (interleukin (IL)-1 receptor-associated kinases 1 and 4) are replaced by kinase-inactive mutants. Components of the TLR7-MyD88-IRAK4-IRAK1 pathway have an important role in driving lupus in other mouse models.4–6 For example, like Tnip1, polymorphisms in Tlr7 predispose to lupus in multiple human populations, suggesting that therapies might be developed by targeting this pathway.7 8
The finding that lupus in ABIN1[D485N] mice is prevented by crossing to knock-in mice expressing a kinase-inactive mutant of IRAK19 suggested that a specific IRAK1 inhibitor may prevent lupus but impair innate immunity less profoundly than a specific IRAK4 inhibitor since the TLR-MyD88-IRAK4-IRAK2 and TLR-MyD88-IRAK4-IRAK3 signalling pathways should remain intact. Many dual inhibitors of IRAK1 and IRAK4 have been developed but to our knowledge, the only IRAK1 inhibitor that inhibits IRAK4 much less potently is pacritinib,10 a drug that was recently approved for myelofibrosis. Interestingly, pacritinib also inhibits the protein tyrosine kinase JAK2, which is required for IL-6 signalling. Crossing ABIN1[D485N] mice to IL-6 KO mice prevented glomerulonephritis and the increase in serum dsDNA antibodies, two of the hallmarks of lupus. We therefore investigated the effects of pacritinib on the lupus phenotype in ABIN1[D485N] mice.
Methods
Administration of pacritinib
WT mice (n=10) and ABIN1[D485N] (n=11) mice aged 6 weeks were fed with R&M3 control diet (Research Diets) or R&M3 diet containing pacritinib at 300 mg/kg food. Since the mice fed on this pacritinib-containing diet lost weight during the first week, the pacritinib was removed from the diet for 1 week and then reincluded at 150 mg/kg when mice were aged 8 weeks and kept on this diet for a further 10 weeks.
Detection of pacritinib in the blood
Blood was collected in MiniCollect EDTA tubes (Grainer). Samples (10 µL) were diluted in 90 µL sterile water and stored at −20°C prior to liquid chromatography with tandem mass spectrometry to determine the pacritinib concentration.
Purification and stimulation of splenic B cells
Splenic single-cell suspensions were washed with 10 mL Phosphate-buffered saline containing 0.5% (w/v) bovine serum albumin and 2 mM EDTA and pelleted by centrifugation (4 min at 450× g). The cells were incubated for 10 min on ice with CD16/CD32 antibody (10 µg/mL) (Biolegend), followed by incubation for 20 min with biotin-labelled anti-mouse CD11c, CD11b, CD3ε, TER119 antibodies (Biolegend) (1 µg per 107 cells). Unbound antibody was washed away, the cells labelled using streptavidin microbeads (Miltenyi Biotech) and passed through LD columns (Miltenyi). The purified splenic B cells were counted, seeded in 12 well plates at a density of 2×106/mL, incubated with pacritinib and stimulated with IL-6 as specified in figure legends. The cells were lysed in 70 µL 1% (w/v) SDS containing a protease inhibitor cocktail (Roche) and benzonase (50 units/mL), subjected to SDS-PAGE, transferred to polyvinylidene difluoride membranes, and immunoblotted with antibodies recognising STAT3 phosphorylated at Tyr705 (Cell Signaling Technology (CST), 9131) or all forms of STAT3 (CST, 4904S).
Other reagents and methods used are detailed elsewhere.4 11 12 Recombinant anti-F4/80 antibody (Abcam, ab111101) was used at 1:100.
Results
Pacritinib was given to ABIN1[D485N] mice at 8 weeks, 4 weeks before the onset of lupus, and disease severity compared with that observed in ABIN1[D485N] and WT mice fed with control diet. The average pacritinib concentration in the blood 10 weeks after feeding the drug was 2.1 µM (female mice) and 1.65 µM (male mice) (online supplemental figure 1A,B). The ABIN1[D485N] mice gained less weight than their WT littermates whether fed on control diet or pacritinib-containing diet (online supplemental figure 2A,B). Towards the end of the study, 3 of 11 ABIN1[D485N] mice fed with the control diet exceeded the 15% weight loss permitted in the study and were culled. In contrast, no ABIN1[D485N] mice fed with pacritinib showed any signs of distress, nor did they lose weight.
Supplemental material
Supplemental material
Pacritinib prevented splenomegaly and the increase in splenic GCB and TFH cell numbers (figure 1A–C). Interestingly, total splenic B cell numbers were similar in WT and ABIN1[D485N] mice fed on control diet, but severely reduced in mice fed with pacritinib (figure 1D), indicating that the drug impacts B cell development.

Effect of pacritinib on spleen size and splenic immune cell populations. WT and ABIN1[D485N] knock-in mice aged 8 weeks were fed for 10 weeks with RM3 control diet or RM3 diet containing 150 mg/kg pacritinib. Spleens were photographed and weighed (A) (the vertical bar=1 cm), and the numbers of germinal centre B (GCB) cells (B), follicular T helper (TFH) cells (C), total B cells (D), neutrophils (E), patrolling monocytes (pMo) (F) and inflammatory monocytes (iMo) (G) were determined by flow cytometry. Each symbol shows the result from a single individual mouse and the exact n numbers are shown in each plot. Statistical significance was calculated using one-way analysis of variance and the Tukey’s post-hoc test or the Kruskal-Wallis and the Mann-Whitney tests. *P<0.05, **p<0.01 and ***p<0.001. n.s., not significant; WT, wild type.
The increase in splenic neutrophil numbers in ABIN1[D485N] mice was abrogated by pacritinib (figure 1E). Increases in patrolling and inflammatory monocytes were prevented partially in the spleen (figure 1F,G) but were unaffected in blood (online supplemental figure 3A, 3B). Neutrophil numbers in the blood of ABIN1[D485N] mice were similar to WT mice and unaffected by treatment with pacritinib (online supplemental figure 3C).
Supplemental material
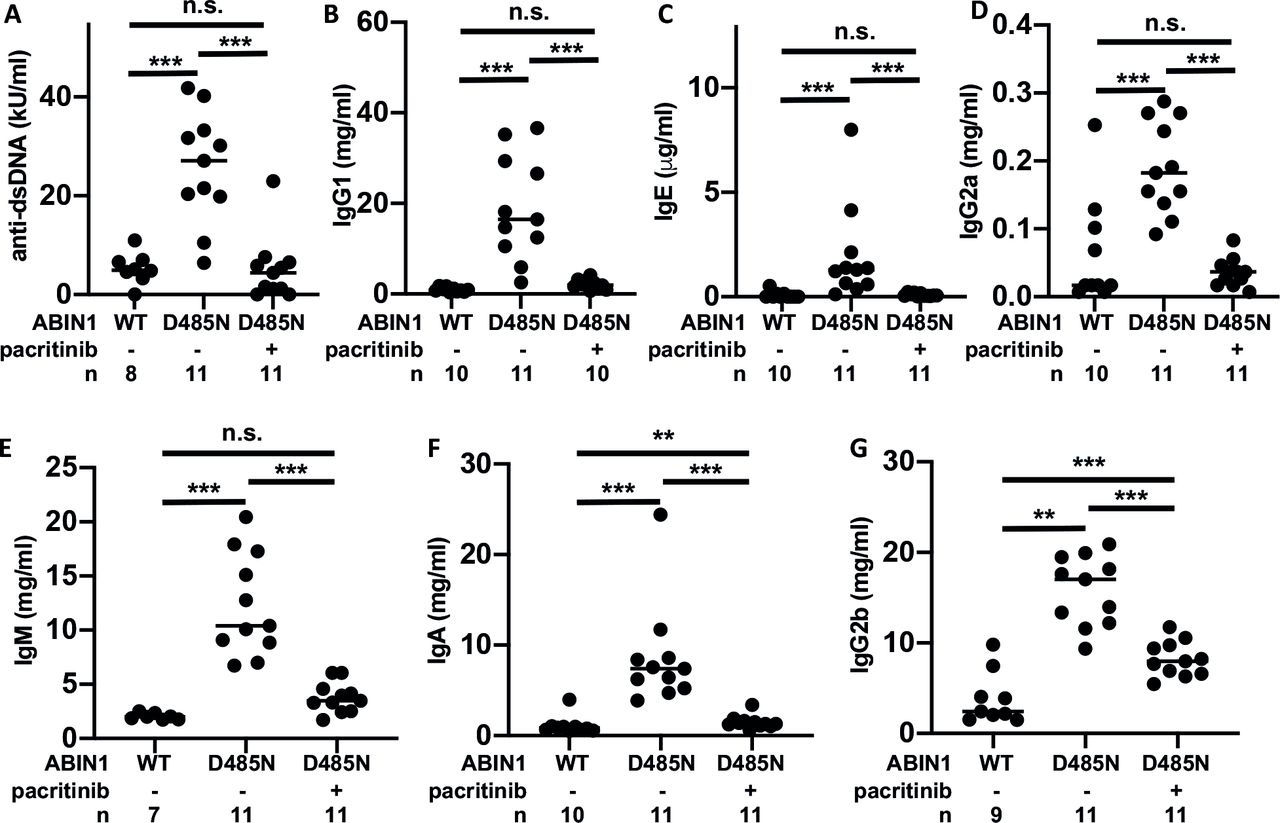
Consistent with the effect of pacritinib on splenic GCB and TFH cells, which are required for immunoglobulin class switching and secretion, the increase in dsDNA antibodies and IgG1, IgE, IgG2a, IgM, and IgA antibodies (figure 2A–F) was prevented by feeding pacritinib to ABIN1[D485N] mice, while the increase in IgG2b was reduced partially (figure 2G). It is possible that the decrease in anti-dsDNA antibodies is a consequence of the reduction in B cell numbers and hence the reduction in immunoglobulin production.

Effect of pacritinib on serum anti-dsDNA autoantibodies and immunoglobulins. As in figure 1, except that blood was collected and serum levels of the following antibodies were measured: (A) anti-dsDNA, (B) IgG1, (C) IgE, (D) IgG2a, (E) IgM, (F) IgA and (G) IgG2b. Each symbol shows the result from a single individual mouse and the exact n numbers are shown in each plot. Statistical significance was calculated using one-way analysis of variance and the Tukey’s post-hoc test or the Kruskal-Wallis and the Mann-Whitney tests. *P<0.05, **p<0.01 and ***p<0.001. dsDNA, double-stranded DNA; n.s., not significant; WT, wild type.
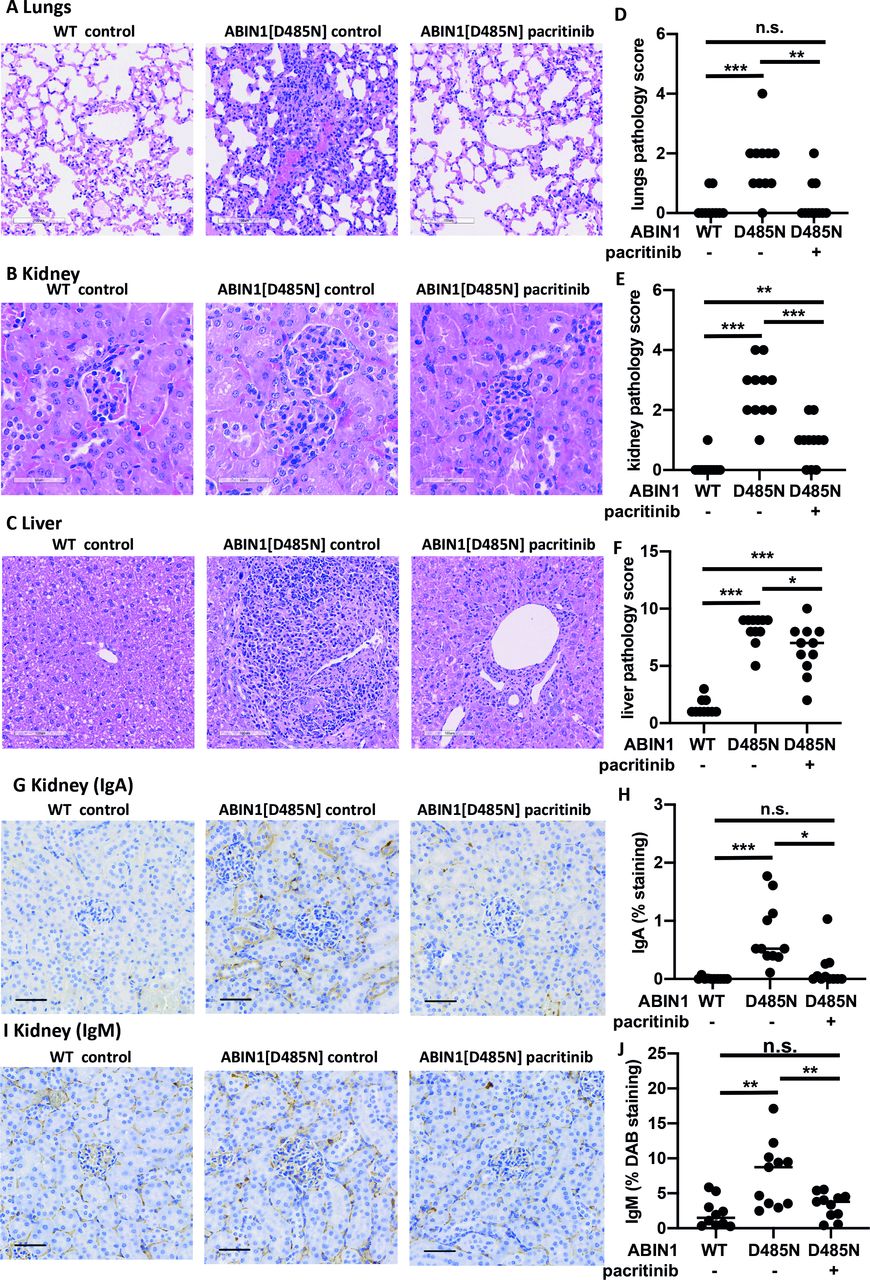
Pacritinib also prevented lung inflammation (figure 3A,D) and reduced kidney (figure 3B,E) and liver inflammation (figure 3C,F) compared with ABIN1[D485N] mice fed on control diet. Staining with anti-PAX5 antibody showed that infiltration of B cells in the lung, kidney and liver was prevented by pacritinib (online supplemental figure 4). CD3+ T cells and F4/80+ cells (macrophages and monocytes) were increased in the kidney and livers of ABIN1[D485N] mice on control diet, but not in the lungs. Interestingly, pacritinib prevented the infiltration of T cells and F4/80+ cells in the kidney, but not in the liver (online supplemental figures 5 and 6). The deposition of immunoglobulins in the kidneys is one of the hallmarks of lupus nephritis. To further investigate the role of pacritinib in preventing kidney pathology, we stained kidney sections with anti-IgA and anti-IgM antibodies.11 IgA and IgM were deposited in the kidneys of ABIN1[D485N] mice fed with control diet but were reduced in ABIN1[D485N] mice fed with the pacritinib-containing diet (figure 3G–J).
Supplemental material
Supplemental material
Supplemental material
{kind=link}
{kind=link}
{kind=link}
Effect of pacritinib on organ inflammation. (A–C) Representative histological images showing H&E staining of the lung (A), kidney (B) and liver (C) of 18-week-old WT (n=10) and ABIN1[D485N] knock-in mice (n=11) after feeding for 10 weeks with RM3 diet with or without 150 mg/kg pacritinib. (D–F) Cumulative pathology scores for the lungs (D), kidney (E) and liver (F) of WT or ABIN1[D485N] mice fed with (+) or without (−) pacritinib. The scoring system is explained in Petrova et al.11 Each symbol shows the result from a single individual mouse. Statistical significance was calculated using the Kruskal-Wallis and the Mann-Whitney tests. *P<0.05, **p<0.01 and *** p<0.001. (G,I) As in B and E, except that representative images of the kidneys stained with haematoxylin (blue) and IgA or IgM antibodies (stained brown with DAB (3,3’-Diaminobenzidine)) (are shown. (H,J) IgA H) and IgM (J) staining was quantified by analysing 10 images per mouse and scored as in Petrova et al.11 The data determined as percentages were logit-transformed and statistical significance was calculated using one-way analysis of variance and the Tukey’s post-hoc test (H,J). *P<0.05 and ***p<0.001. n.s., not significant; WT, wild type.
Discussion
Pacritinib prevented multiple facets of the SLE phenotype in ABIN1[D485N] mice, including splenomegaly, increase in splenic neutrophils, GCB and TFH cells. Moreover, it prevented the increase in many serum immunoglobulins, autoantibodies against self-DNA and the deposition of IgA in the kidney. Some of these effects may be explained by the inhibition of IRAK1 (see the Introduction section). However, pacritinib is also a potent inhibitor of JAK2, which is activated when cytokines, such as IL-6, engage their receptors. As expected, pacritinib partially suppressed the IL-6-stimulated phosphorylation of STAT3 in splenic B cells at a concentration below the serum level reached 10 weeks after administering pacritinib (online supplemental figure 7A). IL-6 plays an important role in GCB cell formation and immunoglobulin class switching,13 suggesting that the effects of pacritinib on these readouts might be explained by inhibition of JAK2 and not IRAK1. However, pacritinib also significantly reduced splenic B cell numbers (figure 1D), which are not reduced in IL-6 KO mice (online supplemental figure 7B). The effect of pacritinib on B cell numbers is therefore independent of IL-6 and might result, for example, from the inhibition of FLT3, a protein tyrosine kinase required for B cell development that is also inhibited by pacritinib.10
Supplemental material
Crossing ABIN1[D485N] mice to IL-6 KO mice prevented autoimmunity but did not affect liver or lung inflammation.4 In contrast, pacritinib completely blocked lung inflammation in ABIN1[D485N] mice and reduced liver pathology to a greater extent than that seen in ABIN1[D485N] mice fed on control diet, indicating that these effects of pacritinib are also not exerted by suppression of IL-6 signalling but by another mechanism, such as inhibition of IRAK1.
Currently, molecular therapies for treating SLE are limited. They include the monoclonal antibodies belimumab and rituximab, which target CD20 on B cells and anifrolumab, which blocks the type 1 interferon receptor.14 15 However, disease manifestation in humans is complex, can affect different organs, and many patients require broad immunosuppression. The present study highlights the potential use of the kinase inhibitor pacritinib for treating lupus in human patients as it has already been approved for another human disease.
Supplemental material
Ethics statements
Patient consent for publication
Ethics approval
Experiments on mice were approved by the University of Dundee Ethical Review Committee under a UK Home Office project licence.
Acknowledgments
We thank Lynn Oxford, Lynn Stevenson and Frazer Bell (Veterinary Diagnostic Services, University of Glasgow) for tissue processing for histological analysis.
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors TP and CF-V performed the experiments and analysed the data. TP and SKN contributed to the experimental design. CS performed histological analysis. PC acquired funding for the study and conceived the study, and PC and TP wrote the paper.
Funding The study was funded by CTI BioPharma Corp.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.