Article Text

Abstract
Objective Friend leukaemia virus integration 1 (Fli-1) regulates chemokine/cytokine expression and thus plays an important role in the development of lupus nephritis. Chemokine CXC ligand 13 (CXCL13) is a chemokine that promotes the formation of ectopic lymphoid structures and has been reported to be associated with the pathogenesis of lupus nephritis. The relationship between Fli-1 and CXCL13 is unknown. This study aims to elucidate whether Fli-1 impacts CXCL13 expression and contributes to the progression of lupus-like nephritis in adult MRL/lpr mouse.
Methods Serum CXCL13 levels were measured in adult wild-type (WT) MRL/lpr mice and Fli-1 heterozygote knockout (Fli-1+/−) MRL/lpr mice (4 months old or older) using ELISA. Renal mRNA expression (CXCL13 and related molecules) was measured using real-time PCR method. Kidneys were removed, stained and evaluated using a pathology scoring system. The grade of CXCL13 or CXC-chemokine receptor type 5 (CXCR5)-positive immune cell infiltration into the kidney was evaluated using immunostaining with anti-CXCL13 or anti-CXCR5 antibodies. We also used immunofluorescence staining with CXCL13- and CD11b-specific antibodies to detect the infiltration of CXCL13/CD11b double-positive immune cells.
Results Serum CXCL13 levels in Fli-1+/− MRL/lpr mice were significantly lower than that in WT MRL/lpr mice (545.5 and 960.5 pg/mL, p=0.02). Renal expression of CXCL13 mRNA and SRY-related HMG box4 (Sox4) (an important factor for B-cell development) levels were significantly lower in Fli-1+/− MRL/lpr mice. Renal histology scores in WT MRL/lpr mice revealed significantly increased glomerular inflammation. Despite similar interstitial immune cell infiltration into the kidney, the number of CXCL13- and CXCR5-positive cells was significantly lower in Fli-1+/− MRL/lpr mice than in WT mice. Furthermore, immunofluorescence staining revealed that Fli-1+/-MRL/lpr mice had significantly fewer CXCL13/CD11b double-positive immune cells.
Conclusion Fli-1 regulates renal Sox4 mRNA expression and infiltration of CXCR5-positive cells as well as CXCL13/CD11b double-positive immune cells into the kidney, which affects CXCL13 expression and lupus-like nephritis.
- Lupus Nephritis
- Chemokines
- Inflammation
Data availability statement
Data are available on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
WHAT IS ALREADY KNOWN ON THIS TOPIC?
The transcription factor friend leukaemia virus integration 1 (Fli-1) played an important role in the pathogenesis of lupus nephritis. However, the relationship between Fli-1 and chemokine CXC ligand 13 (CXCL13) expression has not been investigated.
WHAT THIS STUDY ADDS?
Our findings show that Fli-1 impacts CXCL13 expression in the kidneys of MRL/lpr mice with lupus-like nephritis. Fli-1 heterozygote MRL/lpr mice had significantly lower CXCL13 expression and CXC-chemokine receptor type 5 (CXCR5)-positive immune cell infiltration with lower SRY-related HMG box4 (Sox4) mRNA expression.
HOW THIS STUDY MIGHT AFFECT RESEARCH, PRACTICE OR POLICY
Fli-1 is associated with CXCL13 expression, and inhibiting Fli-1 could be a novel target for treating lupus nephritis.
Introduction
SLE is an autoimmune disease with an unknown aetiology that causes chronic inflammation in multiple organs, including the kidney.1 Lupus nephritis causes diffuse infiltration of inflammatory cells into the kidney, which can lead to renal failure.2 3 Many chemokines and cytokines are involved in the development of SLE and lupus nephritis.4–9 The chemokine CXC ligand 13 (CXCL13), also known as B-cell-attracting chemokine-1 or B-lymphocyte chemoattractant, attracts B cells and promotes the formation of ectopic lymphoid structures.10–14 Follicular dendritic cells (DCs) are the primary source of CXCL13 production in humans; however, epithelial cells, macrophages and T follicular helper (Tfh) cells (but not murine Tfh cells) also produce CXCL13.15 CXC-chemokine receptor type 5 (CXCR5), formerly known as Burkitt’s lymphoma receptor 1 (BLR1), is CXCL13’s sole receptor. CXCR5 is highly expressed in mature B cells and Tfh cells.10 11 Previous research found that the CXCL13/CXCR5 axis was essential for B-cell activation, and CXCL13 is required for lymphoid neogenesis and for the maintenance of secondary lymphoid tissue architecture.16 17 Therefore, the CXCL13/CXCR5 axis is important for regulating B-cell development and homeostasis.14
CXCL13 has been reported to be associated with the pathogenesis of SLE and other autoimmune diseases, including inflammatory bowel disease, multiple sclerosis, rheumatoid arthritis, type 1 diabetes and systemic sclerosis.14 Previous research has suggested that the CXCL13/CXCR5 axis plays a role in the development of SLE and lupus nephritis.18–22 Lee et al have shown that CXCL13 and CXCR5 were found in high concentrations in the renal cortex of patients with lupus nephritis.18 Ishikawa et al discovered increased CXCL13 levels in the kidneys of aged New Zealand Black (NZB)/New Zealand White (NZW) F1 mice. The primary sources of CXCL13 in lupus-prone mice were renal DCs and macrophages.12 22 However, the distinct role of CXCL13/CXCR5 axis in the development of lupus nephritis is still to be clarified.
Friend leukaemia virus integration 1 (Fli-1) is a transcription factor that belongs to the Ets family and binds to DNA sequences that contain a consensus GGA(A/T) motif. Fli-1 is a transcriptional activator or repressor that has been found in endothelial cells, fibroblasts and several haematopoietic lineages.23–26 We previously reported that Fli-1 plays an important role in the development of lupus nephritis.27–29 Zhang et al 27 have reported that heterozygote Fli-1 (Fli-1+/−) MRL/lpr mice (homozygote Fli-1−/− were lethal in utero) showed significantly less renal inflammation and longer survival than wild-type (WT) littermates. Fli-1 regulates cytokine/chemokine expression and immune cell infiltration into the kidney in a lupus model mouse.30–37 Interestingly, peripheral lymphocytes from patients with SLE with active disease exhibited increased expression of Fli-1 transcripts in tandem with disease activity.38 Fli-1 levels were also elevated in patients with newly developed or recurrent lupus nephritis but not in those with SLE who were infected.39 Furthermore, Fli-1 is upregulated in the brains of Alzheimer’s disease and Fli-1 inhibition protects 5xFAD mice (transgenic Alzheimer’s disease mouse model) from pericyte loss and cognitive deficit.40 Taken together, Fli-1 inhibition may present a novel therapeutic target in autoimmune diseases. The relationship between Fli-1 and CXCL13 is currently unknown. This study aims to elucidate whether Fli-1 impacts CXCL13 expression and contributes to the progression of lupus-like nephritis in adult MRL/lpr mice.
Materials and methods
Mice
WT MRL/lpr Fli-1+/+ mice and MRL/lpr Fli-1+/− mice were generated as previously reported.27 Back-crossing with the Fli-1+/− C57BL/6 strain for more than 12 generations produced MRL/lpr Fli-1+/− mice, as described in our previous report. All mice were housed in Fukushima Medical University’s animal institute under pathogen-free conditions. We used aged MRL/lpr Fli-1+/− mice and their WT littermates based on the observation of previous studies; more than half of WT MRL/lpr mice showed fatal outcomes due to the progression of lupus-like nephritis (with high serum double-stranded DNA levels, proteinuria, haematuria, etc) in accordance with ageing, which was prominent in 20–35 weeks old compared with MRL/lpr Fli-1+/− mice.27 28 31 35
Genotyping by PCR
Mice were genotyped using PCR to differentiate between WT and mutant Fli-1 alleles, as previously described.27 Briefly, the PCR primers used include the following: Fli-1 exon IX/forward primer (positions 1156–1180), 50-GACCAACGGGGAGTTCAAAATGACG-30; Fli-1 exon IX/reverse primer (positions 1441–1465), 50-GGAGGATGGGTGAGACGGGACAAAG-30; and Pol II/reverse primer, 50-GGAAGTAGCCGTTATTAGTGGAGAGG-30. DNA was isolated from tail snips of 4-week-old mice using the QIAamp Tissue Kit (Qiagen). The PCR analyses were performed using the C1000 Touch Thermal Cycler (Bio-Rad).
Histopathology, immunohistochemistry and immunofluorescence staining
Kidneys of adult MRL/lpr Fli-1+/− mice and their WT littermates (old mice aged 21–33 weeks) were removed, fixed in formalin and embedded in paraffin. The deparaffinised section was stained with H&E. Light microscopy was used to examine kidney sections, and the grade of kidney inflammation was determined using the previously described pathology scoring system.27 35 Briefly, blinded H&E-stained kidney sections were graded for glomerular inflammation, proliferation, crescent formation and necrosis. Scores ranging from 0 to 3 (0=none, 1=mild, 2=moderate and 3=severe) were assigned to each of these characteristics and then added together to produce a final pathology score. Scores for interstitial inflammation were as follows: 0 for <5% involvement, 1 for 5%–25% involvement, 2 for 25%–50% involvement and 3 for >50% involvement. CXCL13 was detected in paraffin-fixed tissues by anti-CXCL13 antibodies (R&D, Minnesota, USA) and polymer staining reagent using the Histofine Simple Stain Kit (Nichirei, Tokyo, Japan). Similarly, CXCR5+ immune cells were detected using anti-CXCR5 antibodies (R&D). CD11b+ and CXC13+ cells in tissue were detected using fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse CD11b antibodies (BioLegend, California, USA) and CXCL13 antibodies (Abcam) with secondary anti-rabbit IgG conjugated to Alexa Fluor 647 (Abcam).35 CXCL13+ and CXCR5+ immune cell infiltrates were detected as previously described.2 27 Briefly, 10 randomly selected sites were blindly counted to determine the number of CXCL13+, CXCR5+ and CD11b+/CXCL13+ cells per high-power field (magnification 200 or 400 if appropriate); cells were photographed using a microscope equipped with a digital camera (BX63; Olympus, Tokyo, Japan).
Quantitative evaluation of cytokines, chemokines and related cell-signalling molecules by real-time PCR
Transcripts encoding cytokines, chemokines and signalling molecules were evaluated from total RNA extracted from adult mouse renal tissue using the TRIzol reagent (Invitrogen). cDNA was generated using the SuperScript, First-Strand Synthesis System (Bio-Rad). Real-time PCR (RT-PCR) was performed in duplicate using the SYBR Green PCR Master Mix (Thermo Fisher, Waltham, Washington, USA) and two independent RNA preparations, according to the manufacturer’s instructions. The primers for the detection of transcript encoding CXCL13 were acquired from OriGene (Rockville, Maryland, USA), and primers for all other transcripts are listed in table 1. Cycling conditions were as per instructions from the company. RT-PCR was carried out using Real-time PCR Detection System (Applied Biosystems) with relative expression analysis determined by reference to the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH), according to the programme provided by Applied Biosystems. The relative expression of the target genes was calculated using the DDCt method. The RT-PCR expression was 1 for relative quantification in the WT (Fli-1+/+) group.31
Primers used for real-time PCR in this study
Measurement of serum chemokine, cytokine (CXCL13, IL-6) and anti-double-stranded DNA antibodies
Immunoreactive CXCL13 and interleukin 6 (IL-6) in sera from adult mice (26–33 weeks old) were quantified using an ELISA (BioLegend and R&D, respectively), according to the manufacturer’s instructions. Serum anti-dsDNA antibody levels (100 dilutions) were also evaluated using ELISA, in accordance with the manufacturer’s instructions.
Statistics
Results are presented as median±IQR (or mean±SD if appropriate). Quantitative data were analysed using the Student’s t-test with Welch’s correction or Mann-Whitney U test according to the characteristics of data. Correlation analysis between CXCL13 and anti-ds DNA levels was conducted using Spearman’s correlation test. A p-value <0.05 was considered significant. Statistical analyses were conducted using Excel add-ins and Statcel 4 Software (OMS Publishing, Saitama, Japan).
Results
Serum cytokine and chemokine levels
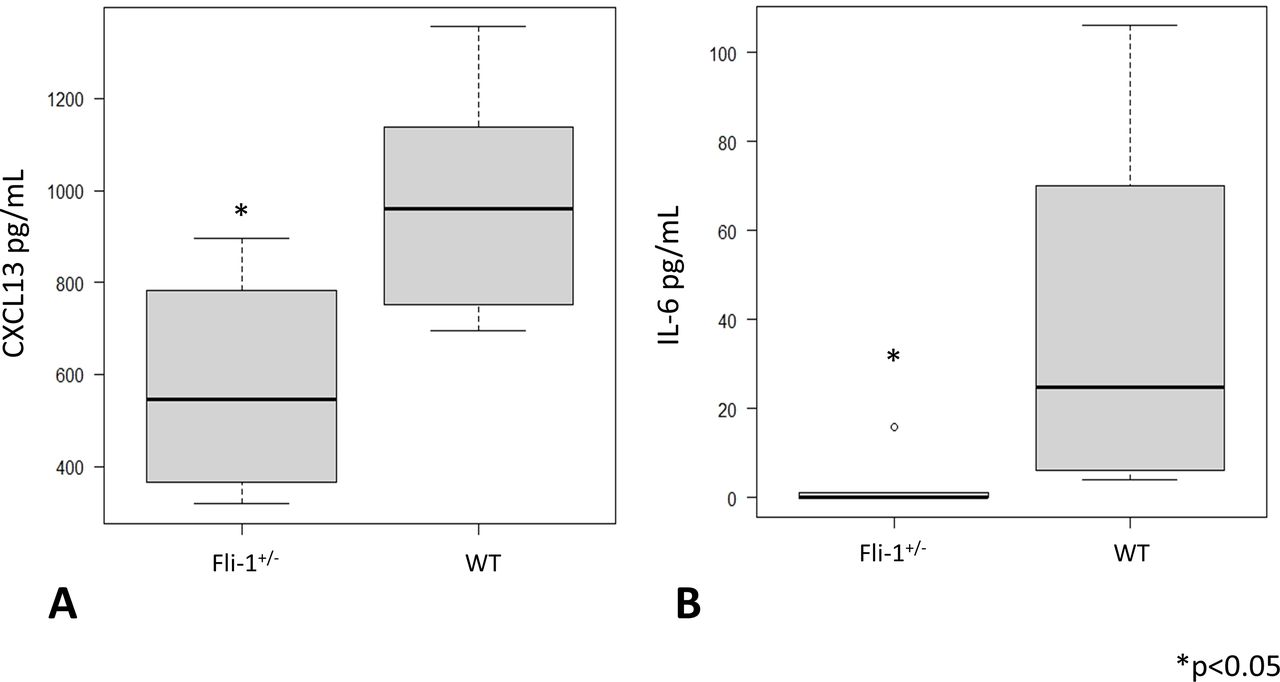
Serum CXCL13 levels and IL-6 levels were measured in MRL/lpr Fli-1+/− mice and compared with age-matched MRL/lpr WT (Fli-1+/+) mice (26–33 weeks old; n=6 each). As shown in figure 1A, median serum CXCL13 levels were significantly lower in MRL/lpr Fli-1+/− mice (545.5 pg/mL, IQR 396.5–736.5) than in WT mice (960.5 pg/mL, IQR 795–1103.5) (p=0.02; figure 1A). Serum IL-6 levels were also decreased in MRL/lpr Fli-1+/− mice (0 pg/mL, IQR 0–0.9) compared with WT littermates (39.5 pg/mL, IQR 10–70) (p=0.01; figure 1B). We also compared serum CXCL13 concentrations between younger (7–9 weeks old) and adult (21–36 weeks old) MRL/lpr mice; as shown in online supplemental figure S2A, younger MRL/lpr mice had significantly lower CXCL13 levels compared with adult MRL/lpr WT mice (p<0.05). The median CXCL13 levels were similar between MRL/lpr Fli-1+/− mice and age-matched MRL/lpr WT (Fli-1+/+) littermates (543.5 pg/mL, IQR 458–629.5 and 539.5 pg/mL, IQR 376.8–644.8, respectively).
Supplemental material
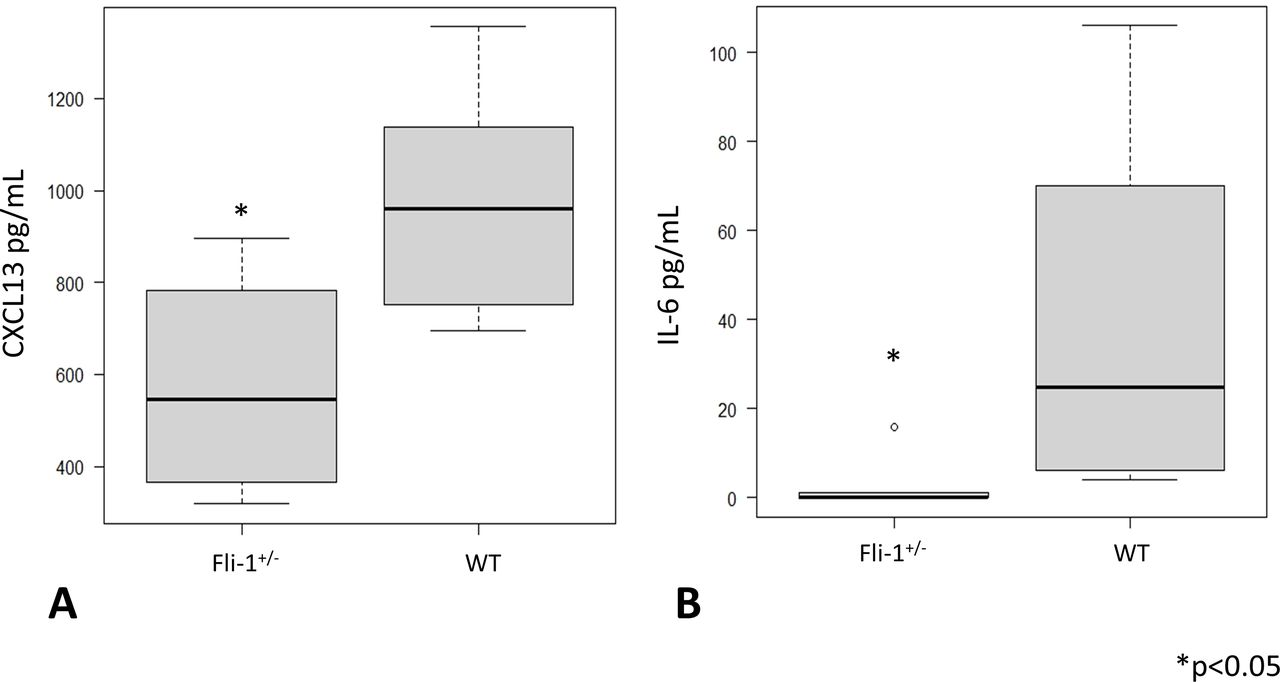
Serum cytokine IL-6 and chemokine CXCL13 levels in adult (26–33 weeks old) MRL WT (Fli-1+/+) and MRL/lpr (Fli-1+/−) mice. (A) Serum CXCL13 and (B) IL-6 levels were significantly lower in MRL/lpr Fli-1+/− mice than in WT MRL/lpr mice (n=6, p<0.05). CXCL13, chemokine CXC ligand 13; Fli-1, friend leukaemia virus integration 1; IL-6, interleukin 16; WT, wild type.
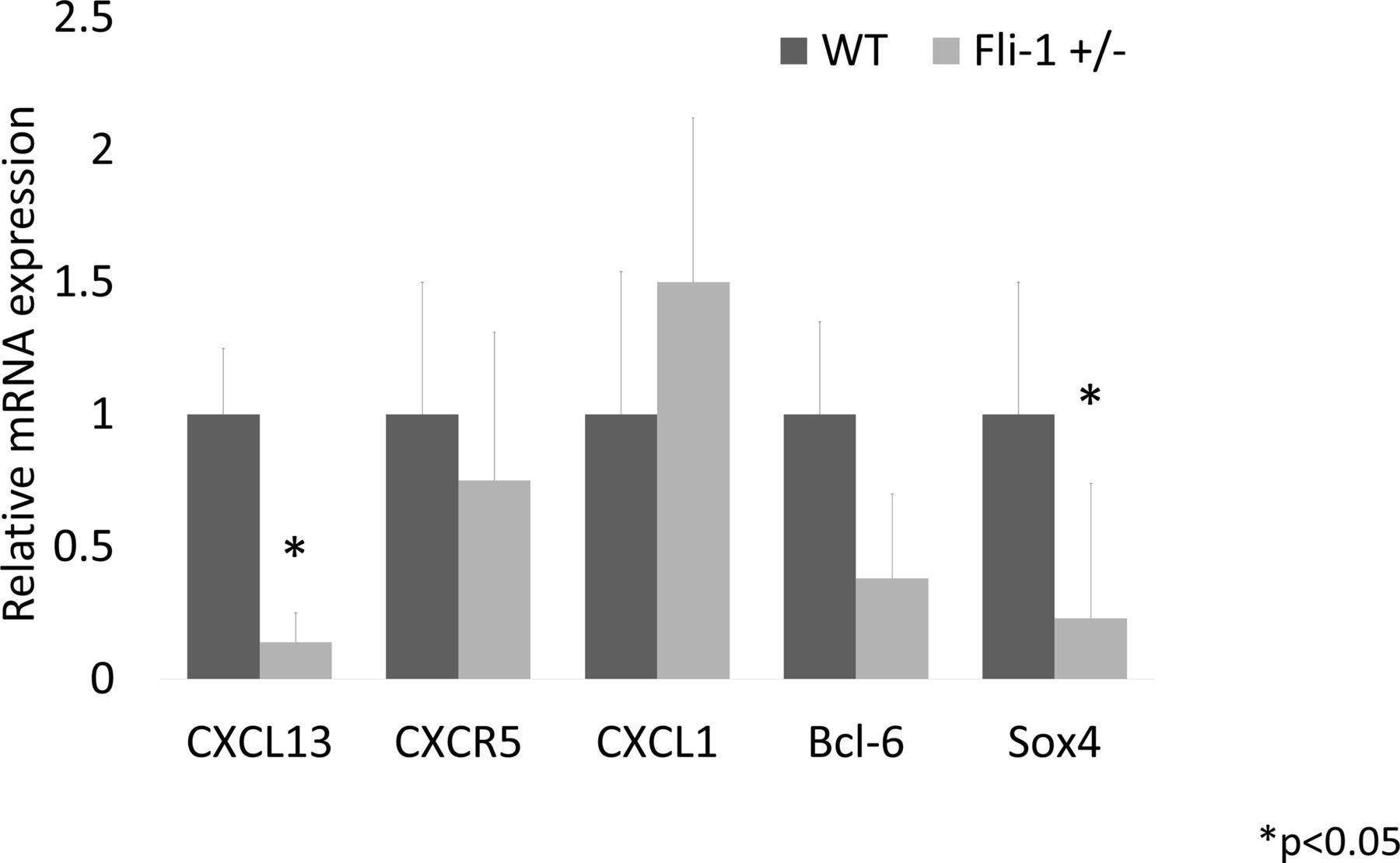
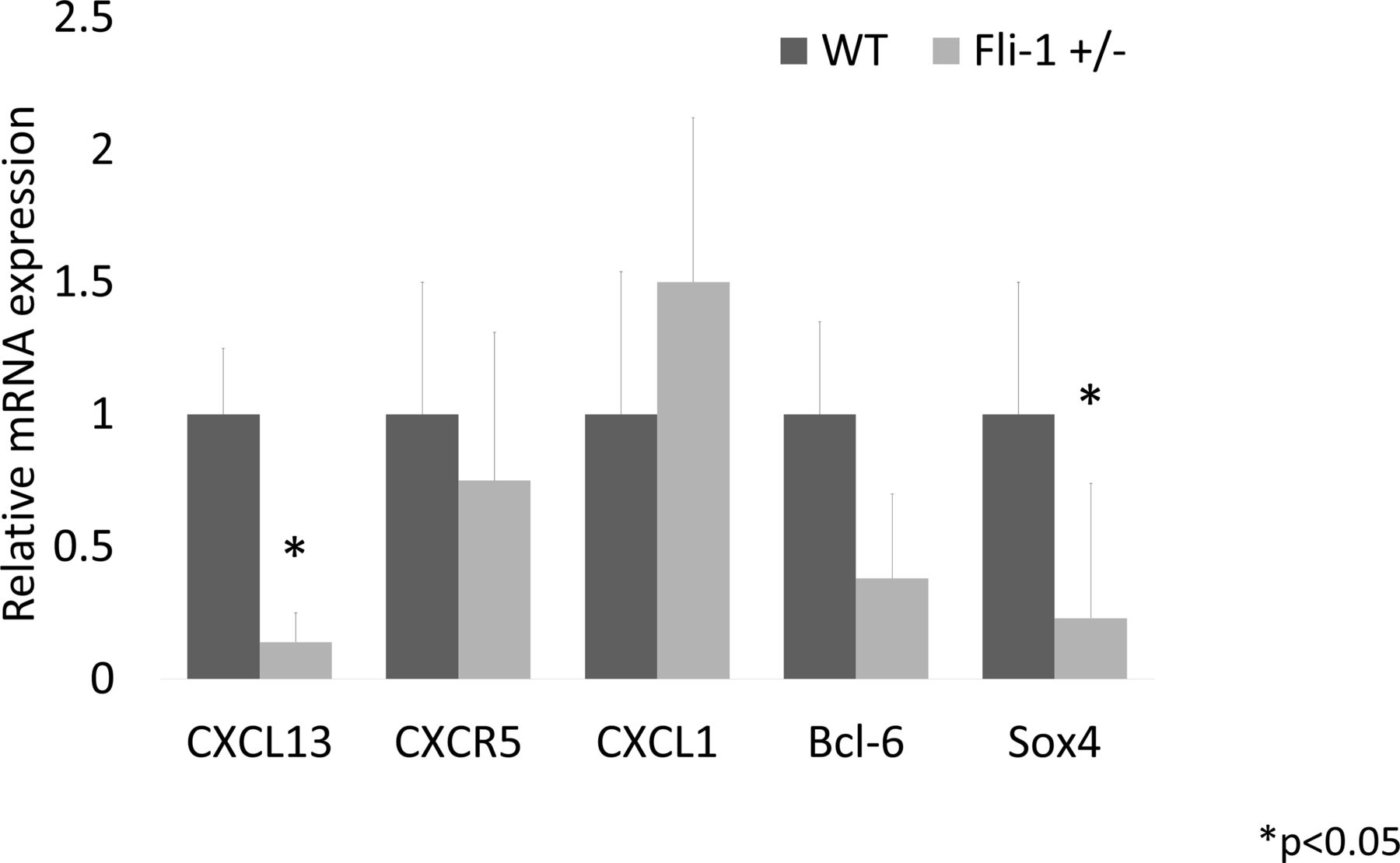
Relative mRNA expression in the kidney of MRL/lpr Fli-1+/− mice compared with MRL/lpr WT control mice (21–33 weeks old, n=4–8). As shown, mRNA expression levels in CXCL13 and Sox4 were significantly lower in MRL/lpr Fli-1+/− mice, whereas CXCR5, CXCL1 and Bcl-6 expression were not significantly lower than the WT control MRL/lpr mice. Bcl-6, B-Cell CLL/Lymphoma 6; CXCL, chemokine CXC ligand; CXCR5, CXC-chemokine receptor type 5; Fli-1, friend leukaemia virus integration 1; Sox4, SRY-related HMG box4; WT, wild type.
Serum anti-dsDNA levels and correlation with cytokine/chemokines
Serum anti-dsDNA levels were measured using ELISA and found that anti-dsDNA levels (100 dilutions) were significantly lower in MRL/lpr Fli-1+/− mice than in WT MRL/lpr mice (p=0.02; online supplemental figure S1A). Interestingly, serum anti-dsDNA levels were positively correlated with serum IL-6 levels (rs=0.58, p=0.03) but not serum CXCL13 (rs=0.24, p=0.22; online supplemental figure S1B,C).
Expression of transcript encoding CXCL13 in the kidney of MRL/lpr Fli-1+/− mice
The mRNA from kidney tissue was isolated and compared between adult MRL/lpr Fli-1+/− mice and control MRL/lpr WT (Fli-1+/+) mice. As a result, the renal relative expression of CXCL13 and SRY-related HMG box4 (Sox4) mRNA levels in Fli-1+/− MRL/lpr mice (21–33 weeks old, n=4–8; figure 2) was significantly reduced. CXCR5 and B-Cell CLL/Lymphoma 6 (Bcl-6) mRNA expression did not differ significantly between the two groups. Unlike adult MRL/lpr mice, CXCR5 mRNA expression in younger Fli-1+/− MRL/lpr mice was significantly reduced compared with WT MRL/lpr mice (7–9 weeks old, n=4; online supplemental figure S2B).
Reduced CXCL13- and CXCR5-positive immune cell infiltrates in MRL/lpr Fli-1+/− mice
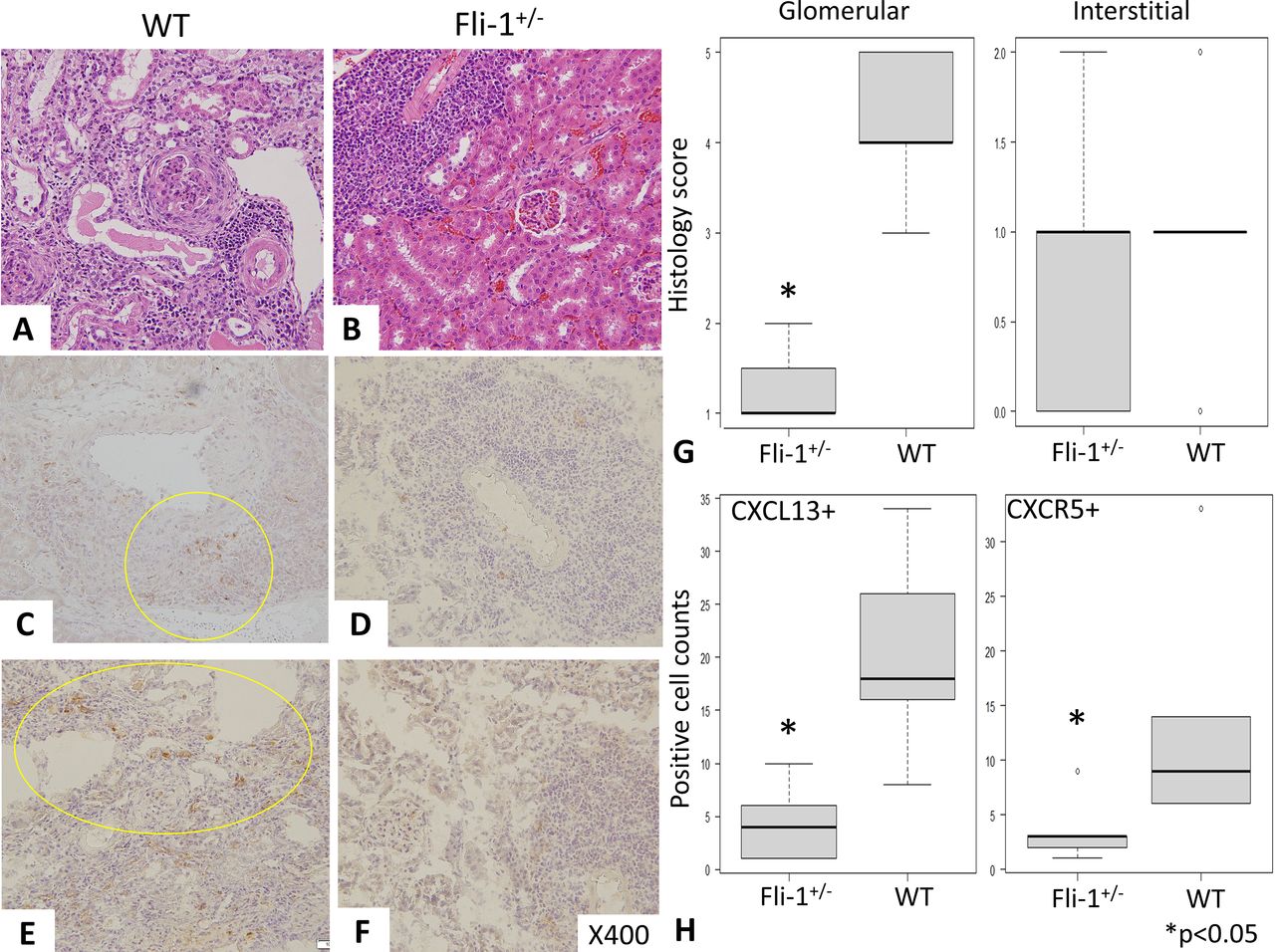
H&E staining revealed severe glomerular inflammation in MRL/lpr WT (Fli-1+/+) mice compared with MRL/lpr Fli-1+/− mice, as shown in figure 3A,B (n=5–7). Renal histology scores showed increased glomerular inflammation in WT MRL/lpr mice (p<0.01), but interstitial immune cell infiltrates into the kidney were similar in the two groups, as shown in figure 3A, B and G. Immunostaining with anti-CXCL13 and anti-CXCR5 antibodies was performed, and the number of CXCL13- and CXCR5-positive cells in MRL/lpr Fli-1+/− mice was significantly lower than that in WT mice (n=5–7, p<0.05; figure 3C–F and H). In contrast, CXCL13 glomerular expression was low in both groups (online supplemental figure S3A,B). Recently, the importance of tertiary lymphoid structure (TLS) lesions has been reported to influence renal prognosis in lupus nephritis; in addition, renal CXCL13 expression has an important role in the infiltration of lymphocytes in TLS.41 42 To investigate this, we have reviewed the infiltration of CXCL13-positive immune cells in the kidney between the two groups. Overall, kidneys of WT MRL/lpr mice showed more intense CXCL13-positive cells not only in TLS lesions (observed around arterioles shown in figure 3C and online supplemental figure S3C) but also in interstitial inflammatory lesions (online supplemental figure S3C).41 In contrast, MRL/lpr Fli-1+/− mice showed smaller number of CXCL13-positive cells in TLS lesions, not spreading surrounding tissues (online supplemental figure S3D).
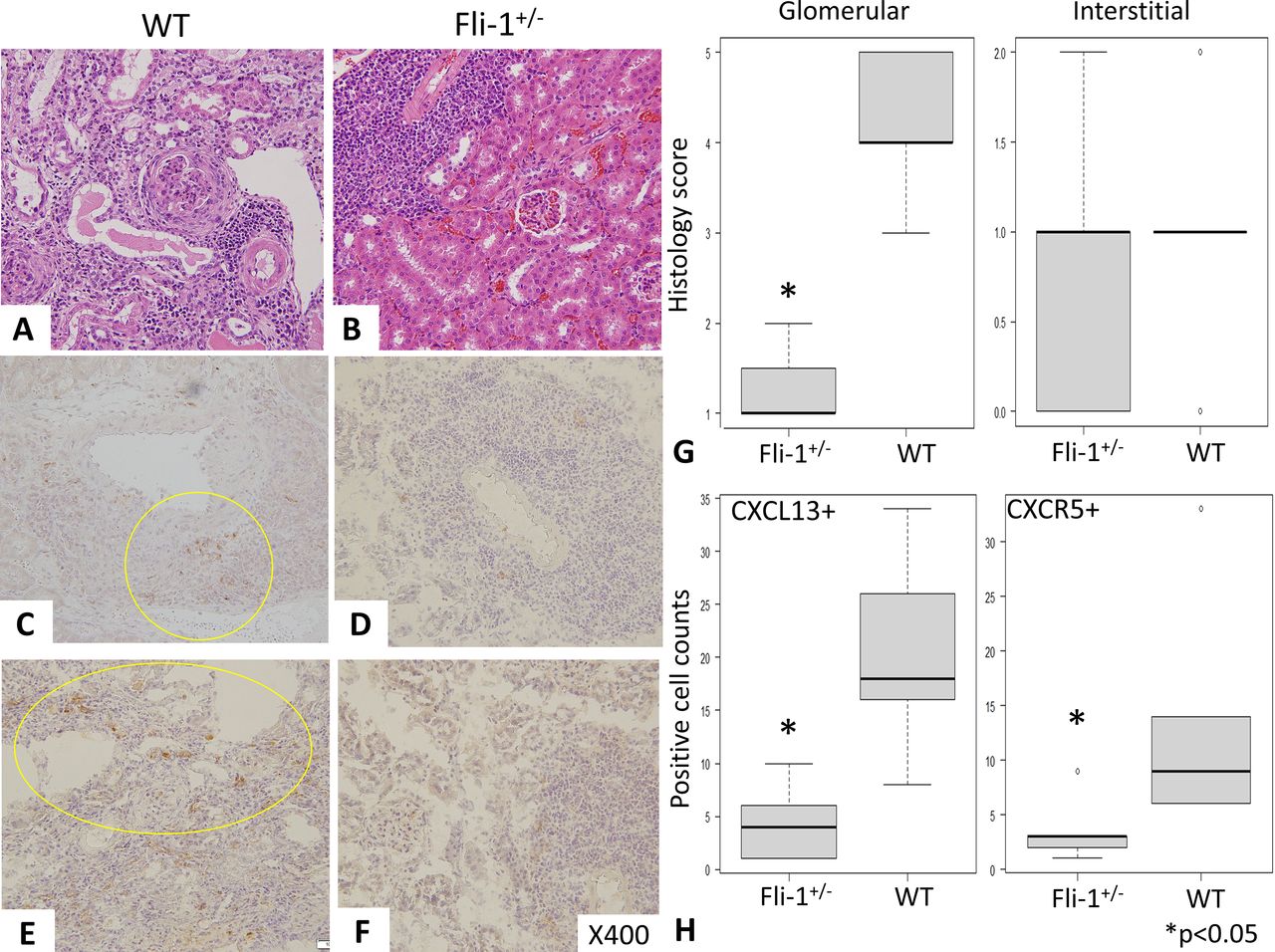
Renal pathology findings (H&E staining and immunostaining) in MRL/lpr wild-type (WT) and MRL/lpr Fli-1+/− mice (21–30 weeks old, n=5–7). (A, B and G) H&E staining showed prominent glomerular inflammation (crescent formation) with higher glomerular scores (Glom) in MRL/lpr WT mice than in MRL/lpr Fli-1+/− mice; in contrast, interstitial inflammation (Int) was similarly observed between the two groups. (C–F) Immunostaining with (C and D) CXCL13 and (E and F) CXCR5 showed decreased positive cell infiltration in the kidney of MRL/lpr Fli-1+/− mice. (H) Pathology scores were significantly decreased in CXCL13- and CXCR5-positive cells in MRL/lpr Fli-1+/− mice. CXCL, chemokine CXC ligand; CXCR5, CXC-chemokine receptor type 5; Fli-1, friend leukaemia virus integration 1.
Detection of CD11b/CXCL13-positive immune cells in the kidney of MRL/lpr WT (Fli-1+/+) and MRL/lpr Fli-1+/− mice
We used immunofluorescence staining with FITC-conjugated anti-CD11b and Alexa Fluor 647-labelled CXCL13 antibodies to detect CXCL13-positive immune cells in the kidneys of MRL/lpr mice. Infiltration of CD11b/CXCL13 double-positive cells was observed in MRL/lpr WT mice (figure 4A, C and E) but not in MRL/lpr Fli-1+/− mice (figure 4B, D and F). Pathology scores also revealed that the number of such cells infiltrating the kidney of MRL/lpr Fli-1+/− mice was significantly reduced (p<0.01, n=5–7; figure 4G). Furthermore, glomerular histology scores were positively correlated with the number of infiltrating CD11b/CXCL13 double-positive cells (rs=0.74, p=0.01) but not with interstitial inflammation (rt=0.38, p=0.21; online supplemental figure S3A,B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Detection of CD11b/CXCL13-positive immune cells in the kidney of MRL/lpr WT (Fli-1+/+) and MRL/lpr Fli-1+/− mice using FITC-conjugated anti-CD11b and Alexa Fluor 647-labelled CXCL13 antibodies. Infiltration of CD11b/CXCL13-positive cells was observed in (A, C and E) MRL/lpr WT mice compared with (B, D and F) MRL/lpr Fli-1+/− mice. (G) The number of infiltration of these cells was significantly decreased in the kidney of MRL/lpr Fli-1 mice (n=5–7, p<0.05). CXCL13, chemokine CXC ligand 13; FITC, fluorescein isothiocyanate; Fli-1, friend leukaemia virus integration 1; WT, wild type.
Discussion
These results show that Fli-1 impacts CXCL13 expression and immune cell infiltration into the kidney of MRL/lpr mice with lupus-like nephritis. CXCL13/CD11b- and CXCR5-positive cells were significantly reduced in MRL/lpr Fli-1+/− mice. Transcripts encoding CXCL13 and Sox4, a transcription factor required for B-cell development, were also downregulated. To the best of our knowledge, this is the first study presenting a relationship between CXCL13 expression and Fli-1. The mechanisms by which Fli-1 impacts CXCL13 expression are still unclear, but they can be explained in a direct and/or indirect manner. Previous studies show that Fli-1 directly regulates many chemokines and cytokines, including monocyte chemoattractant protein-1 (MCP-1), Regulated on Activation, Normal T cell Expressed and Secreted (RANTES), interferon-inducible protein 10 (IP-10), CXCL2, granulocyte-colony stimulating factor (G-SCF), granulocyte-macrophage colony-stimulating factor (GM-CSF), and IL-6.30–34 36 37 Additionally, a recent study found that Fli-1 influenced renal IL-17A expression.35 These chemokines/cytokines are required for the progression of renal inflammation in lupus nephritis.7 8 43 Lema et al 43 have shown that chemokine expression (MCP-1 and RANTES) increases concurrently with mononuclear cell infiltration in a lupus model mouse. In terms of cytokine, IL-17A and IL-6 are important for Th17 cell infiltration and subsequent inflammation in the kidney.35 44 Reduced Fli-1 expression downregulates these molecules, which may alter the milieu of cytokine/chemokine expression in inflamed kidney tissue, ultimately resulting in decreased CXCL13 expression. Many immune cells, including macrophages and DCs, produce CXCL13. The main sources of CXCL13 production in kidney inflammation were macrophages and DCs.12 14 CXCR5 is the CXCL13 receptor that is highly expressed in mature B cells.8 14 Our results showed reduced infiltration of CXCL13/CD11b-positive cells (mainly macrophages) and CXCR5-positive cells (mainly mature B cells). Previous research has shown that the grade of inflammatory cell, CD4+T cell or CD11b-positive cell infiltration into the kidney was slightly lower (but not significantly) in Fli-1+/− MRL mice than in WT littermates (figure 3).35 These findings could imply that Fli-1 impacts cell migration and regulates local inflammation in the absence of similar interstitial immune cell infiltration of kidney tissue. A decrease in CD19-positive cells in Fli-1+/− lupus-prone mice has previously been described, which may be compatible with the present study (decreased renal CXCR5 expression in early timing and CXCR5-positive immune cell infiltration into the kidney, with B cells being the main source of CXCR5-positive cells).18 20 Another possibility is that Fli-1 regulates CXCL13 production directly by binding to the CXCL13 promoter lesion in immune cells/endothelial cells. Because this is an intriguing hypothesis, we are trying to investigate whether Fli-1 directly binds to the promoter lesion that encodes CXCL13. At present, putative Fli-1-binding sites in the promoter lesion of CXCL13 have yet to be described in the literature.
Our result shows that Sox4 mRNA expression is significantly reduced in MRL/lpr Fli-1+/− mice. How did Sox4 deficiency affect inflammation or immune cell infiltration in the kidneys of MRL/lpr mice? Sox4 is a member of the SOX (SRY-related HMG box) gene family, which includes transcription factors involved in cell and organ development and differentiation, as well as the initiation and progression of various cancers, including breast cancer.45 Previous research has shown that the transcription factor Sox4 is required for the survival of pro-B cells in mice.46 On the other hand, Sox4 is involved in transforming growth factor-beta signalling and suppresses the Th2 immune response by regulating GATA3.47 Therefore, skewing the Th2 condition by lowering Sox4 can suppress the Th1 and Th17 immune responses, resulting in lower CXCL13 expression in the kidney. It is also interesting that whether Fli-1 directly regulates Sox4 expression and influences CXCL13 production; however, at present, there are no reports describing the direct regulation of Sox4 via Fli-1. Further investigation is required.
The other possibility is that unrevealed strong CXCL13-producing cells were reduced in MRL/lpr Fli-1+/− mice. In humans, Tfh cells were one of the main sources of CXCL13 production in lymphoid organs; however, murine Tfh cells do not secrete CXCL13.48 On the other hand, a recent study has reported that PD1hiCXCR5-CD4+T cells, called T peripheral helper (Tph) cells, produce CXCL13 in the ectopic lymphoid-like structures in humans, and these cells were regulated by Sox4.49 The existence of Tph cells in lupus-prone mice is unknown; however, unrevealed CXCL13-producing immune cells as mentioned above may be influenced by Fli-1 reduction. Recent reports have shown that TLS in the kidney has an important role in the disease progression of lupus nephritis. Furthermore, CXCL13 expression in the site of TLS is crucial to recruit inflammatory cells in the kidney.41 42 In our observation, the CXCL13-positive immune cells were seen mainly within the immune cell infiltrates in TLS lesions (around arterioles and medulla in the kidney). In addition, the kidney of WT MRL/lpr mice showed wider distribution of CXCL13-positive immune cells, outside of TLS such as periglomerular lesions and kidney interstitium, in contrast to the limited number of CXCL13-positive cells within TLS lesions in MRL/lpr Fli-1+/− mice (online supplemental figure S3C,D). These findings indicate that Fli-1 reduction may impact the infiltration of CXCL13-positive immune cells in TLS lesions and/or surrounding tissues and finally protect kidneys from inflammatory renal damage. Previously, one report has described that glomerular expression of CXCL13 was observed in lupus-prone NZB/NZW F1 mice. However, our results did not show prominent glomerular CXCL13 expression in immunostaining in the kidney of MRL/lpr WT mice (online supplemental figure 3AB). This may imply that inflammatory profiles can be different between NZB/NZW F1 and MRL/lpr mice, even if they show similar phenotypes as lupus-prone mice. Interestingly, serum CXCL13 levels positively correlated with glomerular histology score in our observation (online supplemental figure 4). These facts may indicate that CXCL13 has an important role in the development of lupus-like nephritis in lupus-prone mice. Another previous report described that CXCL13 produced by murine peritoneal macrophages due to Fli-1 deletion contributes to the development of tissue fibrosis, vasculopathy and immune cell activation in systemic sclerosis.50 Patients with systemic sclerosis showed increased serum CXCL13 levels, which were correlated with the modified Rodnan total skin thickness score. This is an intriguing observation: in vitro experiment Fli-1 silencing using small interfering RNA (siRNA) in murine peritoneal macrophages successfully initiated CXCL13 expression on lipopolysaccharide (LPS) stimulation. However, human dermal fibroblasts and microvascular endothelial cells with silencing Fli-1 did not show a significant increase in CXCL13 expression.50 These results may indicate that Fli-1 can act as either repressor or accelerator for the expression of chemokines in each specific cell type (such as peritoneal macrophages and immune cells) during the development of inflammation in autoimmune diseases. Nonetheless, CXCL13 can be an important chemokine for SLE and lupus nephritis: past reports have indicated that serum CXCL13 levels and renal expression of CXCL13/CXCR5 were significantly increased in patients with SLE. Furthermore, CXCL13 levels were positively correlated with the SLE Disease Activity Index in humans.18 19 Blockade of CXCL13 (anti-CXCL13 antibody) attenuates lupus-like nephritis and neuropsychiatric manifestation of MRL/lpr mice.51 52 In any case, further investigation regarding the role of Fli-1 on CXCL13 regulation in autoimmune diseases is needed.
In conclusion, we found a close relationship between Fli-1 and CXCL13 expression in MRL/lpr mice. Reduced Fli-1 expression affects renal CXCL13 expression as well as CXCL13-positive monocytes/macrophages and CXCR5-positive immune cells in inflamed kidneys of lupus-prone mice. Sox4 expression may be important in regulating CXCL13 expression in the kidney. These results also suggest that Fli-1 might be a novel target for agents that treat or control collagen vascular diseases.
Data availability statement
Data are available on reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
All animal experiments were approved by the institutional review board at Fukushima Medical University (no: 2020075). This study was conducted in accordance with the Declaration of Helsinki.
Acknowledgments
We appreciate Enago's assistance with English language editing. We thank Sachiyo Kanno for performing the experiments and providing technical assistance in this study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Presented at Portions of this manuscript were presented at American College of Rheumatology Convergence 2022: https://acrabstracts.org/abstract/transcription-factor-fli-1-impacts-cxcl13-expression-and-renal-inflammation-in-lupus-like-nephritis-of-adult-mrl-lpr-mouse.
Contributors SS, NM and XKZ conceived and designed the experiment. SS, NM, YS, HM, JT and YF were involved in the experiments. SS, KS, SY, YF and TA were involved in the data analysis and writing the original draft. SS, XKZ and KM reviewed and revised the manuscript. All authors have read and agreed to the published version of the manuscript. KM is responsible for the overall content as guarantor.
Funding This study was supported by JSPS Grant-in-Aid for Scientific Research (C) (grant no: 20K08804).
Competing interests None declared.
Patient and public involvement Patients and/or the public were not involved in the design, conduct, reporting or dissemination plans of this research.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.