Article Text

Abstract
Objective MicroRNAs (miRNAs) play an important role in the pathogenesis of various autoimmune diseases including systemic lupus erythematosus (SLE; lupus). We have previously reported a common pattern of miRNA dysregulation in splenic lymphocytes from several mouse models of lupus. In this study, we investigated whether there is a similar miRNAs expression dysregulation in peripheral blood mononuclear cells (PBMCs) and splenocytes in a classical murine lupus model, MRL/lpr.
Method PBMCs were isolated from blood samples of control MRL and lupus MRL/lpr mice aged 14–15 weeks by gradient centrifugation with Histopaque 1083 density media. miRNA TaqMan assays were performed to analyse the expression of 10 lupus-associated miRNAs including miR-182-96-183 cluster, miR-146a, miR-148a, miR-21, miR-31, miR-127, miR-155, and miR-411 in MRL and MRL/lpr PBMCs.
Result In this study, we found that 8 out of 10 examined miRNAs (miR-21, miR-31, miR-127, miR-155, miR-96, miR-182, miR-183 and miR-411) were similarly dysregulated in both PBMCs and splenocytes of MRL/lpr mice when compared with MRL control mice. Only two miRNAs (miR-146a and miR-148a) showed different dysregulation pattern in the PBMCs and splenocytes of MRL/lpr mice. By comparing with the published miRNA data in human lupus, we demonstrated similarity in miRNA dysregulation in murine and human lupus PBMCs.
Conclusion The findings in this study suggest that the miRNA changes observed in PBMCs largely reflect the miRNA dysregulation in cells from the lymphoid organ spleen. Analysis of miRNAs in PBMCs has an advantage over the splenocytes since it allows for monitoring the kinetics of lupus-associated miRNAs expression with peripheral blood cell samples during the development of the disease or after instituting treatment. The similar dysregulation of miRNAs in murine and human lupus PBMCs supports the importance and the feasibility of using murine lupus models to study the pathogenic and therapeutic function of miRNAs in human lupus.
- microRNA
- peripheral blood mononuclear cells (PBMCs)
- splenocytes
- murine lupus model
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0
Statistics from Altmetric.com
Introduction
MicroRNAs (miRNAs) are small, non-coding endogenous RNAs of 18-25 nt in length and are evolutionarily conserved from microorganisms to plants to animals.1 2 The main function of miRNAs is to negatively regulate gene expression at the post-transcriptional phase via binding to the 3′-untranslated region of target mRNAs, leading to mRNA translation inhibition and/or mRNA degradation.1 3 Importantly, miRNAs play key roles in the development of both the innate and adaptive immune system and in the maintenance of immune homeostasis.4–6 Disruption of miRNA biogenesis or function impacts the development and function of different types of immune cells and causes the immune tolerance breakdown and eventually the development of autoimmune-related disorders such as systemic lupus erythematosus (SLE).2 7 8
SLE is a chronic inflammatory autoimmune disease, which causes systemic damage to multiple organs and is characterised by the production of pathogenic autoimmune antibodies, such as antinuclear and antiphospholipid antibodies.9 Recent studies indicated the expression dysregulation of many miRNAs in peripheral blood mononuclear cells (PBMCs) from human patients with lupus when compared with healthy controls.8 The identification of a unique lupus-related miRNAs expression signature in peripheral blood cells and in cell-free body fluid (such as plasma and urine) suggests that miRNAs have the potential to be non-invasive diagnostic biomarkers for human lupus.10–12 Furthermore, these lupus-associated miRNAs could be the targets of new potential therapeutic approaches for lupus treatment and prevention.13
Various murine lupus models have been developed to understand the pathogenesis of human lupus, to characterise the function and pathogenic contribution of specific genes to lupus development and to test different therapeutic approaches for lupus treatment.14 15 To address the fundamental questions of the pathogenic contribution and therapeutic application of specific miRNAs in lupus, the utilisation of specific murine lupus models is instrumental. In our previous study, we have identified a common set of lupus-related miRNAs in the splenocytes from three different spontaneous murine lupus models (MRL/lpr, B6/lpr and NZB/WF1).16 We have further shown that the expression of these lupus-associated miRNAs correlated with the manifestation of lupus and demonstrated sex differences in NZB/WF1 mice.16 17 It is noteworthy that the miRNA profiling studies in human lupus are mostly performed with PBMCs and circulating body fluids. Considering the tissue-specific miRNA expression, it is critical to compare the lupus-related miRNA expression profile from the same cell source in murine lupus and human lupus for translating the data from murine lupus to human lupus. Here, we quantified the expression of 10 selected miRNAs, which have been reported to be highly dysregulated in murine lupus splenocytes and/or human lupus PBMCs, in MRL/lpr lupus PBMCs and compared with control MRL PBMCs.16–23 We further compared the miRNA dysregulation pattern in MRL/lpr lupus PBMCs with previously reported data from human lupus PBMCs.
Materials and methods
Mice
All animal experimental procedures and housing have been approved by the Institutional Animal Care and Use Committee (IACUC) of Virginia Tech. In this study, we used a classical murine lupus model, MRL/MpJ-Faslpr/J (MRL/lpr) mice, which spontaneously develop clinical signs of different autoimmune rheumatic diseases including lupus. The MRL/lpr (stock no: 000485) mice and the control MRL/MpJ mice (MRL, stock no: 00486) were purchased from the Jackson Laboratory (JAX, Maine, USA) and bred in house. All the mice were housed in the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-certified animal facility at the Virginia-Maryland College of Veterinary Medicine, Virginia Tech. The experimental mice were fed with a commercial 7013 NIH-31 odified 6% mouse/rat sterilisable diet (Harlan Laboratory, Madison, Wisconsin, USA) and gave water ad libitum. Given that lupus is a female predominant disease, only female MRL and MRL/lpr mice were used in this study. Carbon dioxide (CO2) was used for euthanasia as required by approved IACUC protocol.
PBMC isolation
The MRL/lpr mice and age-matched MRL control mice aged 14–15 weeks were euthanised by CO2 inhalation. The blood samples were collected from mice immediately following euthanasia through the cardiac puncture. PBMCs were isolated from MRL/lpr and MRL mouse blood samples by gradient centrifugation. Briefly, about 1 mL blood was diluted by an equal volume of phosphate-buffered saline (PBS), loaded into 15 mL tubes containing 3 mL Histopaque 1083 (density=1.083 g/mL, Sigma-Aldrich, Saint Louis, Missouri, USA) and then centrifuged at 400 g for 15 min at room temperature with the brake off. The interphase layer (enriched with PBMCs) was collected and washed with 5 mL PBS. The isolated PBMCs were stained with mouse Percp-Cy5.5-anti-CD11b and PE-anti-Ly6G (eBioscience/Thermo Fisher Scientific, Asheville, North Carolina, USA), then analysed by flow cytometry to determine the efficiency of removal of neutrophils. The aliquots of PBMCs pellets were frozen at −80°C for RNA extraction.
RNA extraction
As we previously reported,17 total RNA containing small RNA was extracted from PBMCs using the miRNeasy Mini Kit (Qiagen) following the manufacturer’s instructions. During RNA extraction, on column DNase digestion was performed to remove any residual DNA in the RNA samples. The concentration and purity of RNA were determined by the NANODROP 2000 spectrophotometer (Thermo Fisher Scientifi). Samples with an OD260/280 ratio at around 2.0 were used for TaqMan miRNA assays.
TaqMan miRNA assay
As we previously reported,16 the TaqMan microRNA assays (Applied Biosystems/Thermo Fisher Scientific) were used to quantify the miRNAs expression by following the manufacturer’s instructions. Briefly, the total RNA samples were reverse-transcribed into cDNA with TaqMan MicroRNA Reverse Transcription Kit, and then followed by quantitative real-time PCR with TaqMan miRNA assay kits using the 7500 Fast Real-Time PCR system (Applied Biosystems). The relative expression level of miRNA was normalised to small RNA control, snoRNA202 and calculated using the equation 2-ΔΔCt (ΔCt=Cttarget miRNA-CtsnoRNA202, ΔΔCt=ΔCtMRL/lpr -ΔCtMRL).
Statistical analysis
Quantitative data are presented in graphs as means±SEM. Statistical differences were analysed with JMP software (Statistical Discovery, Cary, North Carolina, USA). Differences between MRL and MRL/lpr groups were compared by unpaired, two-tail Student’s t-test. p <0.05 was considered to be statistically significant.
Results and discussion
Similar miRNA dysregulation pattern in MRL/lpr lupus PBMCs and splenocytes when compared with control MRL mice
In our previous report, we have identified a common miRNA dysregulation pattern in the splenocytes of different murine lupus models including MRL/lpr, B6/lpr and NZB/WF1 when compared with their respective controls.16 Numerous dysregulated miRNAs have also been identified in human lupus patients and are believed to play a significant pathogenic role in human lupus. The current miRNA profiling data in human lupus was mostly derived from PBMCs, peripheral blood CD4+ T cells and/or plasma. It is not practical to compare the miRNA dysregulation in murine lupus splenocytes with that in human lupus PBMCs, which hinders translating the research results from murine lupus models to human lupus study. To determine whether miRNAs are similarly dysregulated in PBMCs and splenic lymphocytes in MRL/lpr mice, we analysed the expression of 10 previously reported lupus-related miRNAs in MRL/lpr PBMCs and compared with control MRL PBMCs.
As indicated in figure 1, the neutrophils (CD11b+Ly6G+) were significantly depleted from MRL and MRL/lpr whole blood cells following Histopaque 1083 density gradient centrifugation. The neutrophil percentages in the isolated MRL and MRL/lpr PBMCs were less than 1% (0.52±0.08%), while they were about 10%–16% in red blood cell-lysed whole blood samples (12.1%±1.57%).
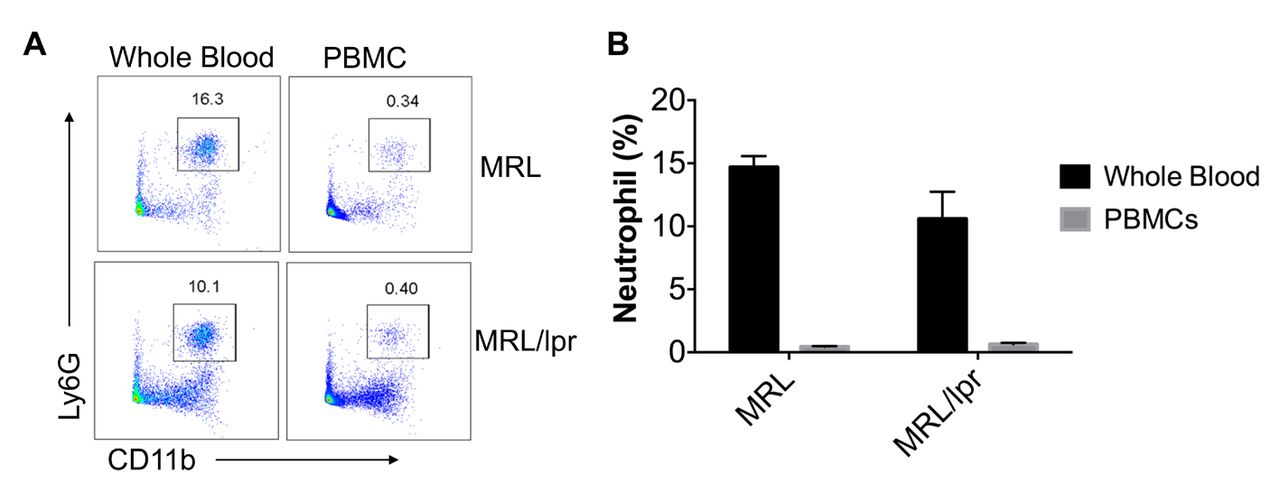
Neutrophils were significantly depleted from isolated PBMCs. (A) Flow cytometry analysis of neutrophils (CD11b+Ly6G+) in whole blood and isolated PBMCs (the box depicts neutrophils). Shown are the representative fluorescence activated cell sorting (FACS) plots from four independent experiments . (B). The summay graph showed the pcercentage (means±SEM) of neutrophils in whole blood and isolated PBMCs (n=4 each). PBMCs, peripheral blood mononuclear cells.
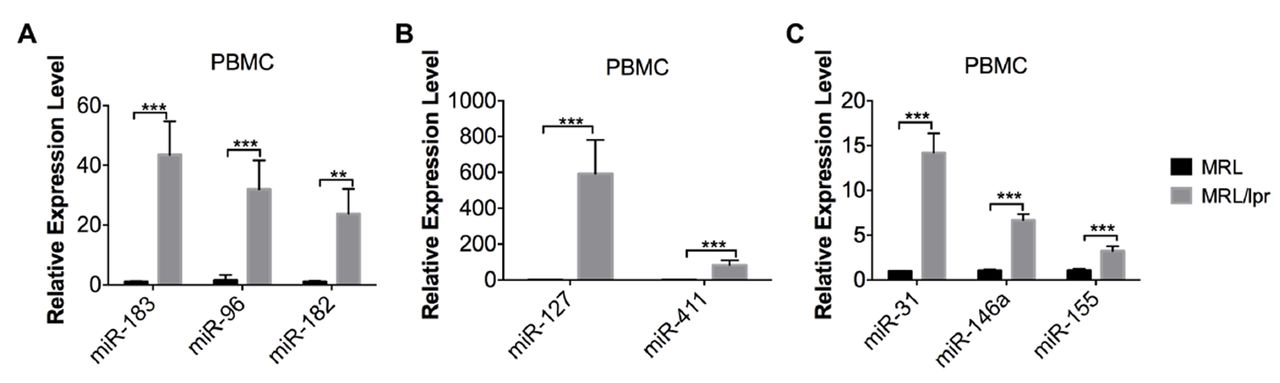
Based on our previous work, the miR-183-96-182 miRNA cluster and genomic imprinted DLK1-Dio3 miRNAs such as miR-127 and miR-411 were substantially upregulated in splenocytes from MRL/lpr mice when compared with that from MRL control mice.16 19 Here, we demonstrated that miR-183-96-182 cluster, miR-127 and miR-411 were also dramatically upregulated in PBMCs from MRL/lpr mice when compared with MRL controls (figure 2A,B). In addition, miR-31 and miR-155 were also upregulated in MRL/lpr lupus PBMCs, mirroring the upregulation observed in MRL/lpr lupus splenocytes (figure 2C).16 In our previous study, we observed a significant upregulation of miR-146a in purified splenic CD4+ T cells but not in whole splenocytes.16 In this study, we observed a significant increase of miR-146a in MRL/lpr lupus PBMCs (figure 2C).
miR-21 and miR-148a have been shown to be upregulated in both MRL/lpr splenic CD4+ T cells and human lupus PMBCs or CD4+ T cells, contributing to lupus pathogenesis by targeting DNA methylation pathway.18 23 Although the miRNA microarray was not sensitive enough to detect the upregulation of miR-21 and miR-148a in MRL/lpr lupus splenocytes in our previous study,16 the miRNA TaqMan assay revealed a significant upregulation of both miR-21 and miR-148a in the splenocytes from MRL/lpr mice when compared with MRL splenocytes (figure 3A). In PBMCs, we observed that miR-21 expression was upregulated, while miR-148a expression was not significantly changed in MRL/lpr mice when compared with that from control MRL mice (figure 3B).
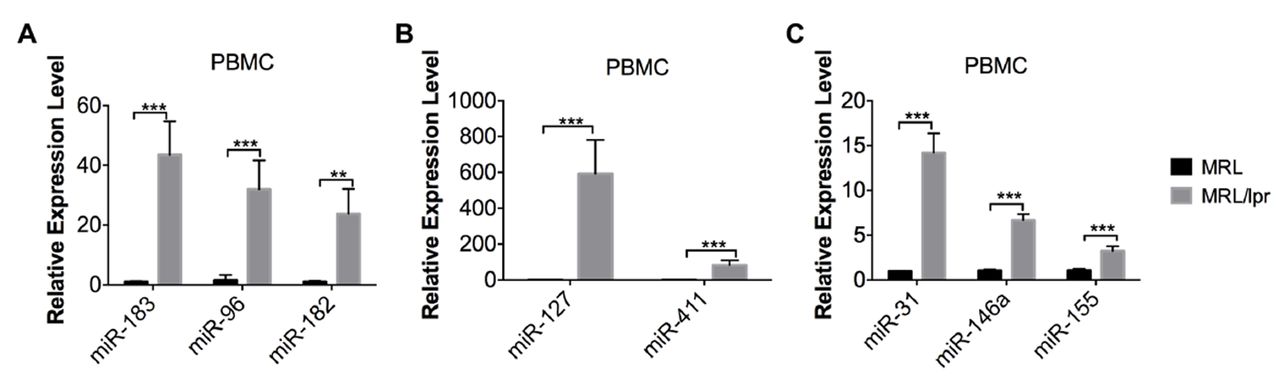
Upregulation of lupus-associated miRNAs in MRL/lpr PBMCs. The miRNA TaqMan analysis showed the upregulation of miR-183-96-182 miRNA cluster (A), genomic imprinted DLK-Dio3 miRNA, miR-127 and miR-411 (B) and other lupus-related miRNAs such as miR-31, miR-146a, and miR-155 in PBMCs from MRL/lpr mice aged 14–15 weeks when compared with the PBMCs from age-matched control MRL mice. Graphs show means±SEM (n=4). Student’s t-tests were performed (MRL vs MRL/lpr). *p<0.05, **p<0.01 and ***p<0.001. PBMCs, peripheral blood mononuclear cells.

{kind=link}
{kind=link}
{kind=link}
Quantification of miR-21 and miR-148a in the splenocytes and PBMCs from MRL and MRL/lpr mice. The miRNA TaqMan analysis of miR-21 and miR-148a in splenocytes (A) and PBMCs (B) from MRL and MRL/lpr mice aged 14–15 weeks. Graphs show means±SEM (n=4). Student’s t-tests were performed (MRL vs MRL/lpr). **p<0.01 and ***p<0.001. PBMCs, peripheral blood mononuclear cells.
Together, our data revealed that 8 out 10 selected lupus-related miRNAs were similarly upregulated in both murine lupus splenocytes and murine lupus PBMCs. The expression of two miRNAs, miR-146a and miR-148a, were differentially altered in the MRL/lpr splenocytes and PBMCs.
Comparison of miRNA dysregulation in human lupus PBMCs and murine lupus PBMCs
miRNA dysregulation in human lupus PBMCs has been extensively reported.8 18 20–23 However, there is no report of miRNA dysregulation in human lupus spleen tissues due to the limitation of obtaining spleen biopsy samples. With the data generated from murine lupus PBMCs in this study, we compared the dysregulation of miRNA in murine and human lupus PBMCs to address the commonality of miRNA dysregulation in human and murine lupus (table 1).
Comparison of miRNA dysregulation in human and MRL/lpr lupus PBMCs, and in MRL/lpr lupus splenocytes
In this study, we demonstrated that most lupus-associated miRNAs are commonly upregulated in PBMCs and splenocytes from MRL/lpr mice (table 1), except that miR-146a is upregulated only in PBMCs and that miR-148a is upregulated only in splenocytes. These data suggested that the miRNA dysregulation pattern in PBMCs largely reflects the miRNA dysregulation in the lymphoid organ spleen. By comparing with the published data from human lupus PBMCs, we found that 6 out of 10 analysed miRNAs are commonly dysregulated in human and murine lupus PBMCs and that only one miRNA, miR-31, was oppositely dysregulated in human and murine lupus PBMCs. As of yet, there is no published data for the three remaining miRNAs (miR-96, miR-183 and miR-411) in human lupus. However, miR-182, which is clustered with miR-96 and miR-183, was upregulated in human lupus PBMCs.18 miR-411 is clustered with miR-127, which was upregulated in human lupus PBMCs,18 at the genomic imprinted DLK1-Dio3 region. We have reported the common upregulation of a cluster of miRNAs at DLK1-Dio3 region in MRL/lpr splenocytes.19 Further investigation with advanced and more sensitive miRNA detection technology is necessary to determine the expression dysregulation of miR-96, miR-183 and miR-411 in human lupus PBMCs.
MiR-31 has been shown to target the linage-specific transcription factor Foxp3 directly, suggesting a potential role of miR-31 in negative regulation of Tregs development and function.24 In a different study, miR-31 was shown to negatively regulate Treg development by targeting retinoic acid-inducible protein 3 (Gprc5a).25 Depletion of miR-31 in CD4+ T cells decreased the severity of experimental autoimmune encephalomyelitis as the result of enhanced development of Tregs.25 MiR-31 was upregulated in the PBMCs of human patients with ankylosing spondylitis, a common inflammatory rheumatic disease,26 and in the target cells of psoriasis, keratinocytes.27 MiR-31 also targeted serine/threonine kinase 40, a negative regulator of NF-κB signalling, leading to enhanced inflammatory cytokine production.27 In addition, Yan et al 28 reported that NF-κB activation induced miR-31 expression, which suppressed protein phosphatase 6 (ppp6c) to promote epidermal hyperplasia in psoriasis. Depletion of miR-31 in keratinocytes decreased acanthosis and reduced the disease severity in the murine model of psoriasis. Together, these studies suggested a potential proinflammatory role of miR-31 by negatively regulating Treg development and promoting NF-κB signalling. Nevertheless, miR-31 was reported to be downregulated in human lupus and contributed to lupus pathogenesis by targeting a different signalling pathway. miR-31 was downregulated in peripheral blood CD4+ T cells and plasma from human patients with lupus, and its expression was negatively correlated with disease severity.29 30 Fan et al further demonstrated that miR-31 was a positive regulator of interleukin (IL)-2 through targeting Ras homologue gene family member A (RhoA). Decreased miR-31 led to upregulation of RhoA, which led to impaired IL-2 promoter activity and defective IL-2 production in lupus T cells.29 Here, we demonstrated that miR-31 was highly upregulated in PBMCs and splenocytes of MRL/lpr mice when compared with control MRL. Further study is needed to understand whether increased miR-31 contributes to lupus pathogenesis in MRL/lpr mice by regulating Tregs and/or NF-κB signalling pathway.
miR-146a plays an important role in the negative regulation of Toll-like receptors-mediated inflammatory responses by targeting TNF receptor-associated factor 6 (TRAF6) and IL-1R-associated kinase (IRAK)-1.31 Depletion of miR-146a led to the development of a spontaneous autoimmune disorder in mice as the result of heightened macrophage inflammatory response.32 Furthermore, miR-146a regulates T cell-mediated inflammation by negatively regulating NF-κB activity and controls Treg-mediated regulation of Th1 responses and inflammation by targeting STAT1.33 34 Depletion of miR-146a specifically in CD4+ T or Tregs cells led to Th1-mediated autoimmunity characterised by the accumulation of activated T cells, overproduction of the Th1 cytokine interferon gamma ( IFNγ) and systemic inflammation.3435 miRNA-146a was downregulated in human lupus PBMCs and played a pathogenic role in lupus by negatively regulating the type I interferon pathway through targeting interferon regulatory factor-5 and signal transducers and activators of transcription-1.22 Nevertheless, the upregulation of miR-146a has been observed in human patients with other chronic autoinflammatory disease such as rheumatoid arthritis (RA) and Sjögren’s syndrome (SS).35 36 miR-146a was upregulated in PBMCs and synovial tissue/fibroblasts from human patients with RA but had no effect on the expression of TRAF6 and IRAK-1. The authors surmised that increased miR-146a in RA was unable to function properly to suppress inflammation, leading to prolonged tumour necrosis factor-alpha production and inflammation in the patients with RA.35 In addition, miR-146a was upregulated in IL-17-producing cells from human patients with RA and correlated with IL-17 expression. Increased miR-146a may contribute to RA pathogenesis by regulating IL-17 production, particular at the early stage of RA.37 Closely related to lupus, SS is also considered as an interferon-driven autoimmune disease. miR-146a was significantly upregulated in the PBMCs and salivary glands, the target tissue of SS, from human patients with SS when compared with healthy controls.36 The upregulation of miR-146a was more dramatically in SS than that observed in RA. There was no correlation between upregulated miR-146a and the expression of type I IFN signature gene MX1 expression in patients with SS. Furthermore, the increased miR-146a and elevated production of inflammatory cytokines (such as IL-1β and IL-18) in the patients with SS contradicted to the negative inflammation regulatory role of miR-146a that was observed from in vitro study.36 The connection between upregulated miR-146a and SS pathogenesis still remains unclear. Although the MRL/lpr model is considered as a classical murine model for lupus, MRL/lpr mice develop RA and SS clinical signs and have also served as the animal model for RA and SS studies.38 39 Therefore, the upregulation of miR-146a in MRL/lpr PBMCs may reflect the development of RA and SS in this model. Reduced miR-146a expression has been reported in PBMCs of two different groups of Chinese patients with lupus.22 40 Nevertheless, a recent study revealed a significant upregulation of miR-146a in the PBMCs of a group of European patients with lupus.18 This variance could be explained by genetic background of lupus patients (Chinese vs European) and also various of other experimental factors.18 Given the important role of miR-146a in the negative regulation of inflammation, further investigation is warranted to fully understand the functional contribution of upregulated miR-146a in the context of autoimmunity.
It is also interesting to observe that although miR-148a was highly upregulated in MRL/lpr splenocytes, it was not significantly changed in MRL/lpr PBMCs. miR-148a was upregulated in PBMCs and CD4+ T cells from human patients with lupus, which targeted DNA methyltransferases (DNMTs) directly to induce DNA hypomethylation in lupus CD4+ T cells.21 23 While we observed an increase of miR-148a expression (figure 3A) and DNA hypomethylation in MRL/lpr splenocytes,19 we did not see a reduction of DNMT1 in MRL/lpr splenocytes when compared with MRL splenocytes (unpublished data). This suggested that miR-148a may regulate autoimmunity in MRL/lpr mice by a different mechanism other than targeting DNMT1 directly. In addition, miR-148a was reported to target gadd45a, Pten and Bcl2l11 and play a critical role in the regulation of B cell tolerance and autoimmunity. Further study is warranted to comprehensively understand the functional significance of miR-148a in the lupus pathogenesis.
In this study, we noticed that only miR-31 was oppositely dysregulated in human and murine lupus PBMCs. This discrepancy in human and murine lupus may be explained by the intrinsic difference in the genetic contribution to human and murine lupus. In addition, we need to consider that many factors such as genetic ethnicity, immunosuppressive therapies, diet and environmental exposure may affect the miRNA expression and cause the discrepancy of miRNA expression data with different human lupus populations. Overall, our study demonstrates a similar miRNA dysregulation pattern in PBMCs and splenocytes from MRL/lpr mice, suggesting the miRNA changes determined in PBMCs could reflect the miRNA dysregulation in cells from the secondary lymphoid organ, which is the spleen. This suggests the practicality of using non-invasive circulating miRNA as biomarkers for monitoring the changes in leukocytes of lymphoid organ during disease development and progression. Importantly, we also demonstrated that most miRNAs are commonly dysregulated in PBMCs of both human and murine MRL/lpr lupus, suggesting the commonality of miRNA dysregulation in human and murine lupus. This supports the importance and the feasibility of using murine lupus models to study the pathogenic function of miRNAs in human lupus.
Acknowledgments
The authors would like to thank Melissa Makris for technical support with flow cytometry analysis and Catharine Cowan for critical editing the manuscript. We also thank all the animal care staff at Virginia-Maryland College of Veterinary Medicine animal facility.
References
Footnotes
Contributors RD and SAA: data acquisition, analysis and interpretation: ZW, BH and RD: manuscript drafting and critical revising: ZW, RD and SAA: final approval of submission: RD and SAA.
Funding This work was supported by Alliance for Lupus Research (Award ID: 219631).
Competing interests None declared.
Patient consent Not required.
Ethics approval Institutional Animal Care and Use Committee of Virginia Tech.
Provenance and peer review Not commissioned; externally peer reviewed.
Data statement No additional data are available.