Article Text

Abstract
Objective Punch biopsy, a standard diagnostic procedure for patients with cutaneous lupus erythematosus (CLE) carries an infection risk, is invasive, uncomfortable and potentially scarring, and impedes patient recruitment in clinical trials. Non-invasive tape sampling is an alternative that could enable serial evaluation of specific lesions. This cross-sectional pilot research study evaluated the use of a non-invasive adhesive tape device to collect messenger RNA (mRNA) from the skin surface of participants with CLE and healthy volunteers (HVs) and investigated its feasibility to detect biologically meaningful differences between samples collected from participants with CLE and samples from HVs.
Methods Affected and unaffected skin tape samples and simultaneous punch biopsies were collected from 10 participants with CLE. Unaffected skin tape and punch biopsies were collected from 10 HVs. Paired samples were tested using quantitative PCR for a candidate immune gene panel and semi-quantitative immunohistochemistry for hallmark CLE proteins.
Results mRNA collected using the tape device was of sufficient quality for amplification of 94 candidate immune genes. Among these, we found an interferon (IFN)-dominant gene cluster that differentiated CLE-affected from HV (23-fold change; p<0.001) and CLE-unaffected skin (sevenfold change; p=0.002), respectively. We found a CLE-associated gene cluster that differentiated CLE-affected from HV (fourfold change; p=0.005) and CLE-unaffected skin (fourfold change; p=0.012), respectively. Spearman’s correlation between per cent area myxovirus 1 protein immunoreactivity and IFN-dominant mRNA gene cluster expression was highly significant (dermis, rho=0.86, p<0.001). In total, skin tape-derived RNA expression comprising both IFN-dominant and CLE-associated gene clusters correlated with per cent area immunoreactivity of some hallmark CLE-associated proteins in punch biopsies from the same lesions.
Conclusions A non-invasive tape RNA collection technique is a potential tool for repeated skin biomarker measures throughout a clinical trial.
- autoimmune diseases
- interferon type I
- lupus erythematosus
- systemic
- pharmacogenetics
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
Interferon-responsive gene expression has been associated with cutaneous lupus erythematosus (CLE) pathogenesis and can be measured in skin biopsies; however, biopsy is relatively invasive and can result in infection or scarring.
Skin tape RNA sampling is an emerging modality to potentially replace invasive skin biopsy; this technique has proven useful to inform on disease-specific gene expression in atopic dermatitis and psoriasis diseases and is in use in the clinic to diagnose melanoma.
What does this study add?
To our knowledge, this is the first use of a non-invasive skin tape method to measure RNA expression in CLE skin.
This study is also the first to identify gene signatures that discriminate between CLE-affected and healthy volunteers’ skin and between CLE-affected and CLE-unaffected skin from tape-derived RNA.
How might this impact on clinical practice or future developments?
This tape RNA method and associated gene signatures may be used as surrogate to punch biopsy to further elucidate CLE natural disease course and/or response to treatment. This novel use of the tape RNA technique will promote easier access to molecular data from CLE skin samples while prioritising patient welfare. While confirmation and validation of this methodology in a large-scale study is needed, it shows promise for use in precision medicine—the right treatment for the right patient.
Introduction
Cutaneous lupus erythematosus (CLE) is a chronic autoimmune disease that may present with or without SLE, and can be divided into acute, subacute and chronic CLE subtypes (including discoid lupus erythematosus [DLE] as a major subtype).1 2 Although the pathogenesis of SLE and CLE are not fully elucidated, genetic risk, environmental triggers and abnormalities in the innate and adaptive immune response have been implicated.3 4
Genetic and clinical observations have highlighted an essential role of type I, type II and type III interferons (IFN-I, IFN-II and IFN-III) in SLE and CLE pathogenesis.5–13 A ‘type I IFN signature’ has emerged as a major risk factor for SLE disease activity.5 14–17 IFN signatures, and myxovirus resistance protein A (MXA), a protein induced by IFN, have been detected in the affected skin of patients with active CLE.18–20 The central role of IFN-I in CLE pathogenesis is further supported by data showing improvement in disease activity in participants with SLE with active skin disease treated with agents targeting IFN-I (anti-IFN-α or anti-IFN-I α/β receptor).21–23 Recently, a humanised monoclonal antibody (BIIB059) targeting blood dendritic cell antigen 2 (BDCA-2), uniquely expressed on plasmacytoid dendritic cells (pDCs)—master regulators of the IFN response— showed following treatment a strong association between reduction in MXA expression in skin and reduction in disease activity and demonstrated efficacy in reducing disease activity in CLE.20 24 25
CLE diagnosis and follow-up is often supported by histological examination requiring a punch biopsy.26 Disadvantages with punch biopsy include pain, localised bleeding, infection and potential scarring.27 Consequently, this technique may impede patient recruitment into clinical trials and decrease patient compliance for follow-up visits. Development of a non-invasive technique for CLE biomarker assessment would offer a significant advantage for patient well-being while permitting monitoring of lesions over time.
Adhesive tape sampling is a non-invasive method that allows recovery of cells comprising and associated with the epidermal stratum corneum and granulosum.28–34 Studies in atopic dermatitis (AD), contact dermatitis, psoriasis and melanoma have used non-invasive tape sampling to investigate disease-specific gene profiles and identify genomic tumour phenotypes and disease-associated transcriptomic endotypes.28 29 33–37 Despite the basal epidermal, junctional and dermal anatomic locations of morphological changes in subacute cutaneous lupus erythematosus (SCLE) and DLE, as keratinocytes express increased levels of IFN-I and IFN-III genes and IFN-inducible genes within CLE lesions13 19 38 39 and local IFN may invoke a hypersensitive response in lupus keratinocytes,40 we hypothesised that expression of specific gene signatures in the epidermis would be detected at the site of skin lesions, and perhaps also in non-adjacent unaffected skin in participants with CLE.
This pilot research study assessed whether an adhesive tape device (https://dermtech.com), which is used in clinical practice for melanoma diagnosis, can recover informative RNA signals that would be specific of lupus disease pathogenesis from the epidermis of participants with DLE or SCLE, as compared and contrasted with healthy volunteers (HVs), and whether it can be used to identify a gene signature that may differentiate affected (A) from unaffected (U) skin. To compare RNA tape gene signature results with a traditional sampling method, from each skin lesion, paired tape RNA and punch biopsy samples were taken. Using punch biopsies, protein immunoreactivity from semi-quantitative immunohistochemistry (IHC) was correlated with gene signature scores from tape RNA.
Materials and methods
Study design
This cross-sectional cohort study was performed at two US study sites. Day 1 assessments included the collection of participants’ characteristics, medical history and a photograph of the taping site for all participants. All participants underwent blood sampling and skin tape harvesting and, for participants who consented, an optional skin biopsy on day 1 (online supplemental figure S1); participants with DLE and SCLE also received a skin examination and disease activity assessment.
Supplemental material
Study assessments
Disease activity for participants with DLE or SCLE was evaluated using Cutaneous Lupus Erythematosus Disease Area and Severity Index (CLASI)41 and a Physician Global Assessment. Procedure-related adverse events (AEs) and serious AEs were monitored and recorded.
Study population
Included participants were aged ≥18 years at the time of informed consent. Participants had to present with active DLE or SCLE skin disease (with or without systemic manifestations of SLE).42 43 Concomitant antimalarial or systemic immunosuppressive therapies were allowed if doses were stable for ≥28 days before the day 1 visit. Prednisone or equivalent maximum dosage was 15 mg/day. High-potency topical steroids and/or other topical agents had to be stopped 7 days before the day 1 visit. HVs had to be in good overall health as determined by the investigator. For further details, see inclusion and exclusion criteria in online supplementary materials.
Biomarker samples
Whole blood and punch biopsy samples were collected from 23 HVs, 9 participants with DLE (1 participant did not consent to a whole blood sample and 1 participant did not consent to a punch biopsy sample) and 1 participant with SCLE. Skin tape samples and punch biopsies were collected from participants with CLE-affected skin (CLE-A) and from HVs; only skin tape samples were collected from CLE-unaffected skin (CLE-U). Further details are in online supplementary materials.
RNA extraction and quantification of gene expression
RNA was extracted from tape using DermTech (La Jolla, California, USA; https://dermtech.com) protocols. Gene expression was quantified by quantitative PCR (qPCR) on the OpenArray platform (Thermo Fisher Scientific, Waltham, Massachusetts, USA). Further details regarding sample quality and gene panels assessments (online supplemental tables S1 and S2) are in online supplementary materials.
Immunohistochemistry and image analysis
IHC for MXA, CD45 and CD303 was conducted on formalin-fixed, paraffin-embedded punch biopsies (online supplemental table S3). Immunoreactive areas were computed using custom-designed algorithms in Visiopharm (Hoersholm, Denmark) software. More details are in online supplementary materials.
Statistical analysis
Descriptive statistics were calculated for demographics and disease characteristics for each group. Individual tape samples were plotted on the first two principal components of log2 expression for all candidate genes. Using the gene expression data generated in the final analysis, candidate genes were grouped into four gene clusters based on the hierarchical clustering of the consensus of 1000 K-means results, after bootstrapping 80% of samples at each iteration (ConsensusClusterPlus R package). Using pooled data generated from interim analyses and final analyses, gene set scores and differences in gene set scores among disease groups were estimated using a linear mixed-effects model, with fixed effects for sample type and random intercepts for genes and samples. P values from multiple comparisons were adjusted using Tukey’s correction. Spearman’s correlations and associated p values were calculated to determine the associations between biomarkers from skin tape-derived RNA and IHC based on skin biopsies.
Additional details regarding statistical methods used for the gene expression analyses are included in the supplementary materials.
Computational biology analysis
Distinct gene clusters from K-means analysis were analysed for biological pathway enrichment using Ingenuity Pathways Analysis (Qiagen, Hamburg, Germany). Pathway enrichment p values were adjusted for multiple hypothesis testing.
Patient and public involvement
No patients or members of the public were involved in the design, or conduct, or reporting, or dissemination plans of this research.
Results
Participant characteristics and safety
This cross-sectional study enrolled 23 HVs. Of these, 10 HVs were age-matched to participants with skin disease and included in the current analysis with 9 participants with active DLE and 1 participant with SCLE; baseline characteristics are presented in table 1. The mean ages for the study groups ranged from 55.7 to 58.0 years. The majority were female in the CLE group. Most participants in the HV, DLE and SCLE groups were white. Three participants with DLE had a concomitant SLE diagnosis. Relevant concomitant medication use was documented. Participants with CLE presented with mild to moderate skin disease activity; CLASI activity score41 ranged from 4 to 19 and Physician Global Assessment score was between 10 and 30. Biopsy sites are shown in online supplemental table S4; for biopsy morphological descriptions, see online supplemental table S5.
Participant characteristics
One participant with DLE experienced two procedure-related AEs: itching at the site of biopsy and minor bleeding at the site of biopsy; both were mild in severity and resolved by day 2. No serious AEs or other AEs were reported during the study.
Tape-derived gene expression
Total RNA yields, pooled from 4 tapes per skin site, for 30 tape RNA samples averaged 42.58 ng (SD, 142.50 ng). No tape samples were removed from analysis based on low RNA yield. Gene expression was measured by qPCR amplification of 102 candidate genes. Eight genes were excluded from statistical analysis because the results did not meet quality control cut-off criteria (see online supplemental table S6 for a list of genes removed), resulting in expression analysis of 94 genes. Additional details are available in the supplementary materials.
To visualise a summary of gene expression of a panel of 94 candidate immune genes (online supplemental table S7) for each participant, we plotted tape-derived gene expression along the first two principal components (PCs; PC1, x-axis; PC2, y-axis) representing the largest variance in the dataset. Each component is a linear combination of 94 immune genes (figure 1A). Compared with samples from CLE-U and HVs, CLE-A were shifted forward on the x-axis of PC1, accounting for 60% of the total variance in the dataset. Within disease groups, CLE-A and CLE-U were separated from one another (figure 1A). HVs were shifted slightly backward along PC1.

Principal component (PC) analysis separates disease groups, and unbiased K-means clustering segregates genes by differential gene expression. (A) PC analysis. Using RNA collected with tape and relative expression of 94 immune genes, participants with CLE are differentiated from HVs. Compared with unaffected skin from participants with CLE and HVs, CLE-affected skin is shifted forward on the x-axis of PC1. (B) K-means clustering of expression for 94 candidate genes. Expression of genes amplified from skin RNA collected with tape was segregated into four clusters. Two of four clusters resulted in differential expression between both CLE-affected skin and HV, and between CLE-affected skin and CLE-unaffected skin: cluster 3, named interferon (IFN) dominant, and cluster 1, named CLE associated. Item consensus (x-axis) represents the percentage of instances that expression of a gene falls within the represented cluster after bootstrapping 1000 times. CLE-A, cutaneous lupus erythematosus-affected skin; CLE-U, cutaneous lupus erythematosus-unaffected skin; HV-U, healthy volunteer-unaffected skin; var, variance.
Expression analysis (K-means) revealed four gene clusters
After determining that tape-derived RNA gene expression from a 94-candidate gene panel could distinguish disease groups, we sought to compile gene clusters with the greatest potential to differentiate between CLE and HV groups. We examined gene expression across all groups using the consensus of 1000 K-means clustering results from bootstrapping. Genes amplified from skin tape-collected RNA were segregated into four clusters, as shown in figure 1B.
We then examined the differential between the mean expression of each cluster in CLE-A versus HV and in CLE-A versus CLE-U (online supplemental table S8). Two of four clusters resulted in differential expression for both comparisons (figure 1B): cluster 3, IFN dominant included 14/18 genes in the cluster that were IFN responsive; and cluster 1, or CLE associated, was composed of a mixture of genes implicated in CLE pathogenesis. The IFN-dominant cluster (cluster 3) differentiated CLE-A from HV (23-fold change; p<0.001) and CLE-U (eightfold change; p=0.002), respectively (figure 2A). The CLE-associated cluster (cluster 1) differentiated CLE-A from HV (fourfold change; p=0.005) and CLE-U (fourfold change; p=0.012), respectively (figure 2B). A third gene cluster (cluster 2) composed of a mixture of IFN-associated and CLE-associated genes (CLE2-associated cluster) also distinguished CLE-A from HV skin (threefold change; p=0.001) but for this cluster, the difference between CLE-A and CLE-U did not reach the level of significance (online supplemental figure S2A and S2B).

Summary scores for interferon-associated and CLE-associated gene clusters differentiate HVs from CLE-A and CLE-A versus CLE-U groups. Gene expression in CLE-affected skin, CLE-unaffected skin and HVs for (A) the interferon-dominant gene cluster (cluster 3) and (B) the CLE-associated gene cluster (cluster 1). Each coloured symbol represents a single participant. CLE-A, cutaneous lupus erythematosus-affected skin; CLE-U, cutaneous lupus erythematosus-unaffected skin; HV-U, healthy volunteer-unaffected skin.
Ingenuity pathway analysis
To explore the relationship between gene clusters found by differential expression between disease groups and their commonly cited pathways in the literature, we used Ingenuity Pathway Analysis (Invitrogen, Carlsbad, California, USA). Analysis of cluster 1 (CLE-associated genes) revealed significant associations with pathways involved in communication between innate and adaptive immune cells, TREM1 signalling and granulocyte adhesion and diapedesis (online supplemental table S9). Analysis of cluster 3 (IFN dominant) revealed associations between genes in the IFN-signalling pathway and the role of hypercytokinaemia/hyperchemokinaemia in the pathogenesis of influenza and pathways associated with pattern recognition receptors that detect bacteria and viruses (online supplemental table S9).
Heat map of gene-level expression
To examine the contribution of individual genes to clusters 1 and 3, we developed a heat map of gene-level expression relative to the mean of the HV samples. The greatest individual gene log2 fold change above the mean of the HV was visualised in the CLE-A disease group versus the HV group in the IFN gene cluster (range, 3.1–5.9; figure 3). Unsurprisingly, genes with the strongest differential between CLE-A and the mean of the HV were IFI6, IFI44, MX1 and USP18 (range, 5.5–5.9), all of which are IFN response genes. A strong differential was also seen between CLE-A and CLE-U in the IFN cluster. There was a less robust differential between CLE-A and the mean of the HV in the CLE-associated cluster (range, 1.4–3.8), for example, ISG15, GZMB and ITGAX with log2 fold changes ranging from 3.3 to 3.8, and a weak differential between CLE-A and CLE-U in the CLE-associated cluster.
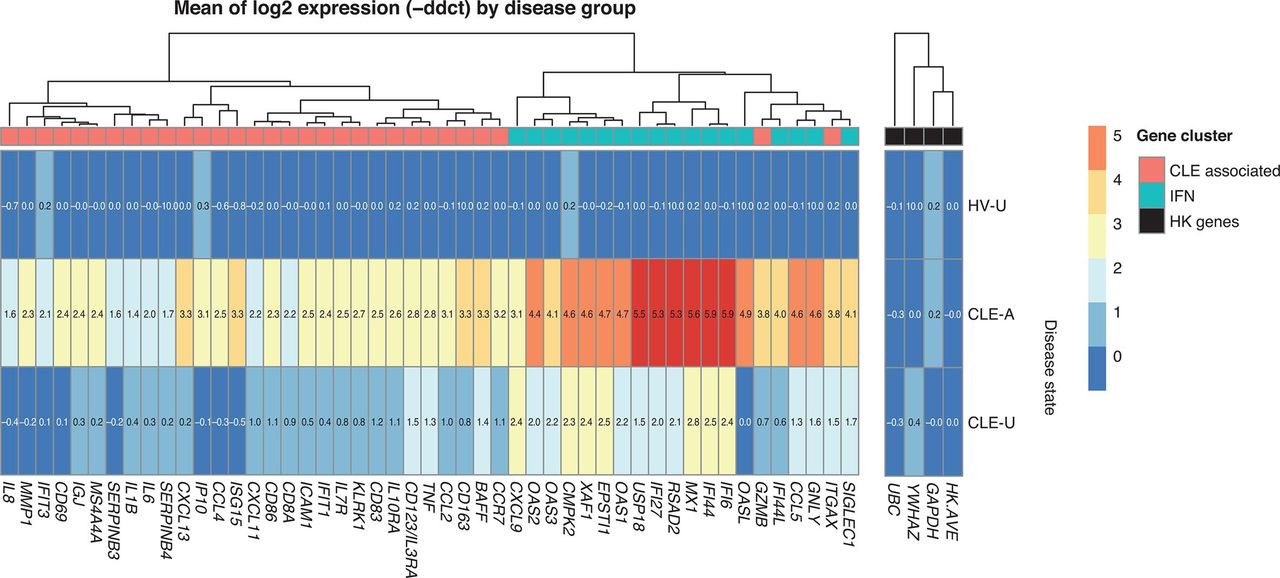
Heat map of interferon (IFN)-associated and CLE-associated gene expression by disease group. Log2 fold change from the mean of HV for each gene in the IFN-associated (blue boxes in top row) and CLE-associated (red boxes in top row) clusters. Log2 fold change for three housekeeping (HK) genes (GAPDH, UBC, YWHAZ), and the average log2 fold change for these three HK genes (HK.AVE), are shown (black boxes in top row). Log2 fold changes are given by colour (red, increased; dark blue, decreased) and by value in each cell. Relatedness of gene expression log2 fold changes is depicted with a dendrogram across the top of the figure. Disease groups are listed along the y-axis. Genes are listed along the x-axis. CLE-A, cutaneous lupus erythematosus-affected skin; CLE-U, cutaneous lupus erythematosus-unaffected skin; HV-U, healthy volunteer-unaffected skin.
Differential expression of interferon genes in skin versus blood
Given the robust differential IFN gene signature in CLE skin, we explored whether expression of IFN genes in blood would discriminate between participants with CLE and HVs. For this analysis, we used an abbreviated 22-gene IFN signature20 to measure the gene expression in paired blood-derived and skin tape-derived RNA samples from five participants with CLE (online supplemental table S10).
Mean change in IFN gene expression between CLE and HV was sevenfold (p=0.016) in whole blood RNA and ninefold (p=0.002) in skin tape-derived RNA (online supplemental figure S3). Mean change in gene expression between CLE-A and CLE-U in skin tape-derived RNA was fourfold (p=0.010; online supplemental figure S3). These data indicate that in patients presenting with active CLE skin lesions, skin tape-derived RNA may be as sensitive—perhaps even more sensitive—than blood RNA for the assessment of IFN gene signature expression.
Correlation of myxovirus resistance protein A, CD45 and CD303 protein quantification
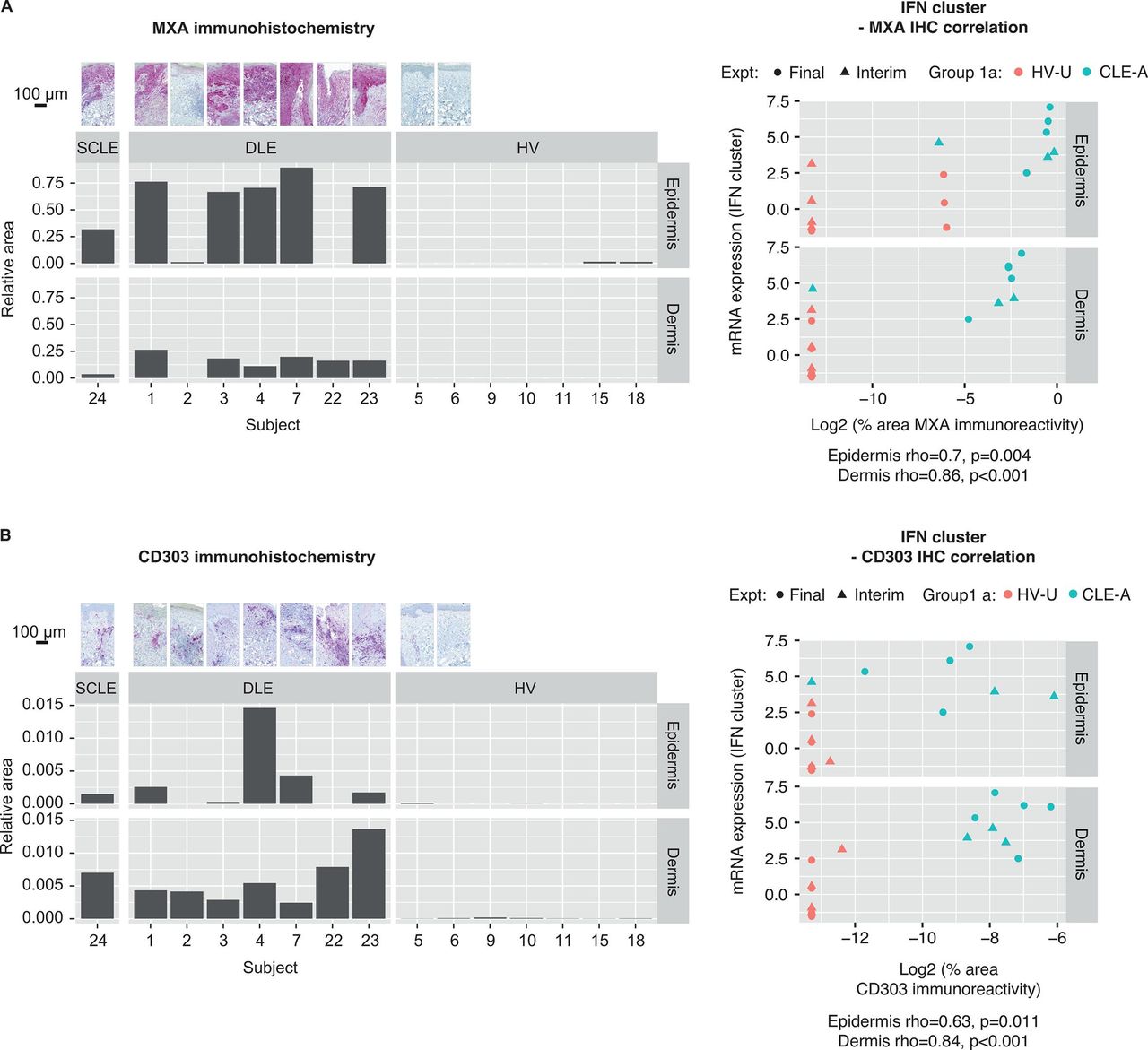
As the traditional method of assessing biomarkers in CLE lesions is punch biopsy followed by IHC, we selected three tissue-based protein biomarkers of CLE activity for comparison with IFN-associated and CLE-associated gene clusters from skin tape-derived RNA. We performed IHC on biopsies taken at the same site as the CLE-A skin tape-derived RNA samples for the following: MXA (annotated as MX1 in RNA clusters), an IFN modulated protein; CD45, for inflammatory infiltrates and CD303 (BDCA-2), a receptor uniquely expressed on pDCs. On images derived from IHC slides, we annotated epidermis and dermis, and compared immunoreactive tissue area for each biomarker by disease group (figure 4, online supplemental figure S4A).
As expected, the MXA immunoreactive area was large and diffusely distributed throughout the epidermis in participants with CLE, with less robust multifocal immunoreactivity in the dermis. MXA expression ranged from very low to absent in HVs. A Spearman’s correlation between per cent area of MXA immunoreactivity and IFN mRNA gene cluster expression was highly significant (dermis, rho=0.86, p<0.001; figure 4A). As most of the IFN gene cluster comprised IFN response genes, including MXA, and MXA is expressed in the epidermis, this result was expected and indicated that tape-derived RNA offers a potential surrogate for MXA protein quantification in punch biopsies.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
An interferon (IFN)-dominant gene signature is correlated with a hallmark IFN-response protein (A) myxovirus resistance protein A (MXA) and (B) CD303+ plasmacytoid dendritic cells demonstrated by immunoreactivity in punch biopsies. Representative photomicrographs of immunohistochemistry (IHC) for MXA and CD303, bar graphs with their quantification and Spearman’s correlations between each IHC biomarker and the IFN RNA summary score. CLE-A, cutaneous lupus erythematosus-affected skin; DLE, discoid lupus erythematosus; Expt, experiment; HV, healthy volunteer; HV-U, healthy volunteer-unaffected skin; mRNA, messenger RNA; SCLE, subacute cutaneous lupus erythematosus.
The CD45 immunoreactive area was most robust in the dermis, with a multifocal distribution, found in participants with CLE and at very low levels in HV skin. Per cent CD45 immunoreactive area correlated with IFN mRNA gene cluster expression (dermis, rho=0.74, p=0.002; online supplemental figure S4B). Across disease groups there was enough of a dynamic range in CD45 immunoreactivity and IFN gene expression to provide a positive correlation between these biomarkers.
CD303 immunoreactive area was greatest in participants with CLE, with a multifocal distribution in dermis tracking along with inflammatory infiltrates. CD303 was very low in HVs. Per cent area of CD303 immunoreactivity strongly correlated with IFN mRNA gene cluster expression (dermis, rho=0.84, p<0.001; figure 4B).
Using the CLE-associated gene cluster, correlations with MXA and CD45 immunoreactive areas were also statistically significant (online supplemental figure S5).
Discussion
This pilot study was designed to assess if a tape device (https://dermtech.com) used in the clinic for melanoma diagnosis36 37 could identify gene signatures relevant to CLE pathogenesis and be used as a tool to monitor CLE biomarkers and potentially replace punch biopsy in the clinical research setting. Using skin tape mRNA collection, two gene clusters were identified with differential expression in CLE-A versus CLE-U and HV skin. Gene expression in these two clusters, IFN-associated and CLE-associated, correlated with MXA, CD45 and CD303, and with MXA and CD45, respectively, protein in punch biopsies by immunoreactive tissue area.
In this study, candidate genes were selected based on their association with CLE pathogenesis, and were enriched with IFN pathway genes likely to distinguish between CLE-A and CLE-U skin. As expected, IFN response genes were plentiful in clusters 1 and 3, the gene clusters with the strongest differential in mean expression between CLE-A and HV, and between CLE-A and CLE-U lesions.
Our finding that IFN pathway genes recovered by tape harvesting distinguish between CLE-A and CLE-U or HV skin is supported in the CLE transcriptomics literature.44–50 Many of the IFN response genes with highest ranking fold changes in previous studies are also differentially regulated in this tape-derived RNA study. These include IFI27, IFI44L, IFI44, OAS1 (blood from participants with CLE),44CCL5, CXCL9, Mx1, OAS2 and AIM2 (affected skin biopsies from participants with CLE)45 46 50 and Mx1, IFI6, OAS2, XAF1, CXCL10 and IFI27 (follicular epithelium from participants with CLE).47
Previous evidence also underpins an important role for cytotoxic NK cell-mediated and T cell-mediated apoptosis in CLE pathogenesis.19 45 51 In support of this hypothesis, in CLE transcript profiling studies comparing CLE lesions with unaffected skin or with HV skin using full-thickness skin biopsy45 or follicular epithelium from plucked hairs,47 upregulated IFN response genes included the apoptosis-related XAF1, OAS1 and OAS2, consistent with our findings. Also convergent with our findings, in previous CLE transcriptome studies, genes expressed in cytotoxic NK and T cells were upregulated, including FAS, GZMB, PRF1 and GNLY.45 The presence of vacuolar degeneration along the basal epidermis due to degenerating and apoptotic keratinocytes is a hallmark of CLE histological diagnosis, highlighting a prominent role for apoptosis and cell-mediated keratinocyte killing in the interface dermatitis characteristic of CLE.
This pilot study was designed to evaluate a tape device as a potential tool in clinical trials to monitor CLE biomarkers that are relevant to CLE pathogenesis. Although limited by a small sample size, results indicated that material of sufficient quality and quantity was obtained to provide relevant information on IFN-associated or CLE-associated gene signature detection. However, additional study is warranted to confirm sensitivity and/or specificity of the skin tape device for use in large efficacy clinical trials in CLE. Previous experience of this skin tape RNA device (Dermtech, La Jolla, California, USA) in the diagnosis of melanoma demonstrated 91% sensitivity and 69% specificity. This device is currently used in clinical practice to confirm the diagnosis of melanoma.36 37
Although differential RNA expression from cytotoxic NK-cell and T-cell genes was observed in this tape RNA study, we cannot rule out the possibility that blood contamination of skin tape-derived RNA samples may have introduced RNA from circulating cells.
We noted that the fold change in IFN pathway genes is more pronounced in skin tape samples than in whole blood. This finding is supported by earlier work, in which after unsupervised clustering, skin CLE samples separated out from blood CLE samples along the first three PCs.48 Additionally, specific Kyoto Encyclopaedia of Genes and Genomes pathways were enriched in the skin compared with blood.48
There are two, non-mutually exclusive mechanisms to explain the finding that mean fold change of IFN genes was higher in skin relative to whole blood involving either pDCs and associated inflammatory infiltrates and/or keratinocytes. In subjects with SLE, pDC numbers are decreased in blood.11 By contrast, pDC numbers are increased in target organs including kidney52 53 and skin.54 55 pDCs isolated from CLE skin biopsies express multiple IFN-α subtypes, IFN-β and IFN-ε.56 As potent producers of IFN, localised, tissue-based enrichment of pDCs as part of an inflammatory infiltrate also expressing elevated levels of IFN-induced genes19 57 may contribute to enhancement of IFN-stimulated gene expression in the target organ. In addition, cross-talk among the cells involved in CLE dermatitis, including NK, B and T, myeloid and pDCs may promote increased expression of IFN by pDCs and keratinocytes.19 58–60
Keratinocytes produce type I IFNs including IFN-κ,39 important in the potentiation of epidermal IFN production in CLE skin lesions.38 In addition to IFN-I, keratinocytes produce type III IFNs.13 In an IHC study, epidermal immunoreactivity for IFN-λ and the IFN-λ receptor was increased in SCLE and DLE lesions versus lesions from other inflammatory skin diseases and healthy participants.13 As an additional source of IFN-λ, it has been shown in a study with ex vivo peripheral blood mononuclear cell cultures that TLR9 ligand (CpG-A)-induced IFN-λ secretion is decreased on treatment with BIIB059,20 which binds to BDCA2,61 a receptor unique to pDCs. In keratinocytes, both IFN-κ and IFN-λ signal via their receptors through the Jak/STAT pathway in common with IFN-α, resulting in overlapping IFN gene response signatures, so both IFN-I and IFN-III could contribute to the differential gene expression in CLE-affected skin versus blood reported here.62
Related to the above mechanisms leading to localised differential expression of IFN genes, we observed increased gene expression of both IFN-associated and CLE-associated gene clusters in affected skin compared with unaffected skin. In unaffected skin, IFN-associated and CLE-associated gene expression was not at the same level as HV, rather the expression level was intermediate between affected skin and HV skin, similar to what has been reported in AD33 63 and psoriasis.34 The finding of an intermediate level of IFN-associated and CLE-associated gene expression in unaffected CLE skin may indicate that there is a low level of inflammation not discernible by gross morphological change.
To our knowledge, no publications describing a comparison between tape-derived and biopsy-derived gene expression are available in CLE. Such studies are published in AD and psoriasis. The two modalities yielded distinct yet overlapping results. Kim et al showed that expression of terminal epidermal differentiation markers in tape-derived HV and AD samples correlated with expression of the same markers in adjacent biopsy.32 Dyjack et al found that in non-lesional AD skin, there was more overlap between skin tape and biopsied epidermis than between either skin tape and biopsied dermis or biopsied epidermis and biopsied dermis.33 Highlighting the relevance of tape sampling for inflammatory skin disease, in an AD type 2 high endotype the most highly differentially expressed genes detected by skin tape were related to activation of TH2 and DC pathways.33 He et al34 found that tape-derived transcriptomes analyses from patients with AD and psoriasis were correlated with a previously published transcriptome biopsy dataset. Interestingly, tape transcriptomes from AD non-lesional skin showed greater fold changes than biopsy specimens for both AD and psoriasis, with tape RNA showing significant upregulation of TH2 products (CCL17/TARC and TNFRSF4/OX40) not found in biopsy specimens.34 Taken together, these data indicate that tape-derived RNA yields distinct and overlapping results relative to biopsy-derived RNA, and certain biomarkers can be assessed by tape RNA in better resolution than biopsy-derived RNA.
In this pilot study, we demonstrate the ability of IFN-associated and CLE-associated gene clusters to distinguish between CLE-A and CLE-U, and HV skin. These data indicate that tape-derived RNA is a potential tool to monitor biomarkers in longitudinal CLE studies, although a study with a larger sample size is warranted to confirm the validity of this approach in CLE.
Acknowledgments
The authors would like to thank the following for their valuable contributions: Zuxu Yao and Talisha Allen from DermTech, Inc., Robert Dunstan, Huo Li, David Martin, Norman Allaire, Marian Themeles, Anne Ranger, Amy Kao, Kejie Li and Margaret Choy Merola. The authors wrote the first draft of the manuscript and Linda Wagner of Excel Scientific Solutions provided editorial support. Miranda Dixon from Excel Scientific Solutions copyedited and styled the manuscript.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
NF and TLR are joint senior authors.
Contributors Conceptualisation: CB, TLR, NF, JFM, DR. Data curation: CGW, TLR, SH. Formal analysis: CGW, WW, TLR. Funding acquisition: NF. Investigation: TLR, CL, JFM, AT, SH, XZ. Methodology: CGW, WW, TLR, GM, XZ. Project administration: CM, CB, TLR, GM. Software: CGW, SH. Supervision: CB, TLR, NF. Validation: TLR. Visualisation: CGW, WW, CB, TLR. Writing—original draft preparation: CB, WW, TLR, JFM. Writing—review and editing: CGW, WW, CB, TLR, CL, NF, JFM, AT, GM, DR, XZ, SH, CR, CM.
Funding Biogen sponsored this study. Biogen provided funding for medical writing support in the development of this manuscript. Biogen reviewed and provided feedback on the article to the authors.
Competing interests JFM is a consultant and/or investigator for AbbVie, Aclaris, Almirall, Avotres, Biogen, Celgene, Dermavant, EMD Serono, Incyte, Janssen, Leo, Lilly, Merck, Novartis, Pfizer, Sanofi Regeneron, Sun and UCB. WW, SH, AT, CM, GM, CB, NF and TLR are employees and shareholders of Biogen. CGW, XZ, CR and DR are former employees of Biogen. XZ, CR and DR are now employees of Vertex, Genomics Institute of the Novartis Research Foundation and Rheos Medicines, respectively. CL has been an investigator for Biogen. NF serves on the board of OMass Therapeutics.
Patient consent for publication Not required.
Ethics approval This study was conducted in accordance with the Declaration of Helsinki and other ethical principles described in Title 45, United States Code of Federal Regulations Part 46, Subparts A and B, and approved by the institutional review boards at Brigham and Women’s Hospital (approval ID # 2013P002615/MGH) and Boston University School of Medicine (approval ID # H-34177). Written informed consent was obtained from all participants. The study started on 6 May 2014 and completed on 26 May 2016.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Requests for data supporting this manuscript should be submitted to the Biogen Clinical Data Request Portal (http://clinicalresearch.biogen.com).
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.