Article Text

Abstract
Objective To evaluate the effects of targeting Ikaros and Aiolos by cereblon modulator iberdomide on the activation and differentiation of B-cells from patients with systemic lupus erythematosus (SLE).
Methods CD19+ B-cells isolated from the peripheral blood of patients with SLE (n=41) were cultured with TLR7 ligand resiquimod ±IFNα together with iberdomide or control from day 0 (n=16). Additionally, in vitro B-cell differentiation was induced by stimulation with IL-2/IL-10/IL-15/CD40L/resiquimod with iberdomide or control, given at day 0 or at day 4. At day 5, immunoglobulins were measured by ELISA and cells analysed by flow cytometry. RNA-Seq was performed on fluorescence-activated cell-sorted CD27-IgD+ naïve-B-cells and CD20lowCD27+CD38+ plasmablasts to investigate the transcriptional consequences of iberdomide.
Results Iberdomide significantly inhibited the TLR7 and IFNα-mediated production of immunoglobulins from SLE B-cells and the production of antinuclear antibodies as well as significantly reducing the number of CD27+CD38+ plasmablasts (0.3±0.18, vehicle 1.01±0.56, p=0.011) and CD138+ plasma cells (0.12±0.06, vehicle 0.28±0.02, p=0.03). Additionally, treatment with iberdomide from day 0 significantly inhibited the differentiation of SLE B-cells into plasmablasts (6.4±13.5 vs vehicle 34.9±20.1, p=0.013) and antibody production. When given at later stages of differentiation, iberdomide did not affect the numbers of plasmablasts or the production of antibodies; however, it induced a significant modulation of gene expression involving IKZF1 and IKZF3 transcriptional programmes in both naïve B-cells and plasmablasts (400 and 461 differentially modulated genes, respectively, false discovery rate<0.05).
Conclusion These results demonstrate the relevance of Ikaros and Aiolos as therapeutic targets in SLE due to their ability to modulate B cell activation and differentiation downstream of TLR7.
- lupus erythematosus
- systemic
- B-lymphocytes
- autoimmune diseases
This is an open access article distributed in accordance with the Creative Commons Attribution 4.0 Unported (CC BY 4.0) license, which permits others to copy, redistribute, remix, transform and build upon this work for any purpose, provided the original work is properly cited, a link to the licence is given, and indication of whether changes were made. See: https://creativecommons.org/licenses/by/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
The transcription factors Ikaros and Aiolos, which are critical for B cell differentiation, are implicated in systemic lupus erythematosus (SLE) pathogenesis.
Targeting Ikaros and Aiolos using the cereblon modulator iberdomide has been proposed as a promising therapeutic agent.
What does this study add?
Targeting Ikaros and Aiolos with iberdomide inhibits the TLR7-mediated differentiation of plasmablasts in human SLE.
Iberdomide inhibits TLR7 and interferon driven autoantibody production by SLE autoreactive B cells.
RNA-sequencing was used to detect differentially expressed genes in iberdomide-treated differentiated SLE plasmablasts, thus identifying genes which are most directly under the control of Ikaros and Aiolos.
How might this impact on clinical practice or future developments?
This study provides strong evidence that therapeutic targeting of Ikaros and Aiolos can ameliorate key pathogenic processes in human SLE.
Introduction
The Ikaros family of zinc-fingers (IKZF) proteins are transcription factors critical for the development of key haemopoietic stem cell lineages and their maturation into effector cells.1 IKZF1 (Ikaros) is an essential regulator of common lymphoid progenitor (CLP) stem cells. Ikzf1-deficient mice lack CLP and fail to generate mature B/T lymphocytes, natural killer and dendritic cells.2 3 IKZF1 also acts as a checkpoint at the pro-B cell to pre-B cell transition stage.4 5 In contrast, IKZF3 (Aiolos) is necessary for memory B cell and plasma cell formation and Ikzf3 deficient mice lack both of these mature B cell populations.6 7
IKZF1, IKZF3 and more recently IKZF2 (Helios) have been identified as susceptibility loci in systemic lupus erythematosus (SLE) in large-scale genome-wide associated studies.8–13 IKZF1 polymorphism rs4917014 was identified as a trans-expression quantitative trait locus, driving upregulation of type 1 IFN genes and downregulation of complement genes.14 Ikaros has also been shown to influence TLR7 signalling,15 representing another link with SLE pathogenesis.16
Ikaros and Aiolos have emerged as the therapeutic targets of the immunomodulatory drugs thalidomide and analogues such as lenalidomide, which act as agonists for the ubiquitination E3 ligase complex cereblon,17 thus inducing ubiquitination and proteasomal degradation of Ikaros and Aiolos.18–20 Thalidomide and its analogues are well established as treatment options for multiple myeloma.21 Lenalidomide inhibits plasma cells differentiation from healthy donors in vitro22 and has been found to be effective in cases of treatment-refractory cutaneous manifestations of SLE.23–25 More recently, a novel cereblon ligand has been developed, iberdomide, which binds with higher affinity than thalidomide and other analogues, resulting in greater Ikaros/Aiolos degradation.26
In preliminary studies, iberdomide reduced Ikaros and Aiolos expression in SLE B cells, while inhibiting antibody production and differentiation of B cells into plasmablasts.27 Iberdomide has undergone a phase I study, which explored the effects of oral administration in healthy volunteers and confirmed the reduction of Ikaros and Aiolos at protein level in B cells, T cells and monocytes.28 Iberdomide also inhibited anti-dsDNA and antiphospholipid antibody production ex vivo from SLE mononuclear cells. Iberdomide reduced absolute B cell counts, augmented IL-2 production from T cells and inhibited IL-1β production in response to proinflammatory stimuli, underpinning its further development for the treatment of SLE, with an ongoing phase II trial (Clinical-Trials.gov Identifier: NCT02185040).
In this study, we set out to investigate the effects of targeting Ikaros and Aiolos with iberdomide on peripheral blood B cells from patients with SLE, specifically on (i) TLR7 and IFN mediated B cell activation and differentiation, (ii) plasmablast/plasma cell differentiation and (iii) transcriptional programmes underlying B cell differentiation.
Materials and methods
Patient recruitment
Peripheral blood and clinical demographics were collected from patients with a diagnosis of SLE from Barts Health NHS Trust and Kings College Hospital NHS Trust. All study participants provided written informed consent at time of sample collection and fulfilled the 2012 updated American College of Rheumatology classification criteria ACR criteria for SLE.29 Patient characteristics are summarised in table 1. Sample collection from patients and subsequent analysis were approved by UK local ethical committee (REC reference 17/WS/0172). Patients or the public were not involved in the design, conduct, reporting or dissemination plans of the research.
Summary of patient characteristics (n=41)
Blood processing and B cell isolation
Peripheral blood mononuclear cells were isolated from peripheral blood using gradient separation with Lymphoprep (Alere), with RBC lysis via incubation in ammonium chloride (RBC-lysis buffer, Stemcell) for 5 min on ice. CD19+ cells were isolated via negative magnetic selection with EasySep Human B Cell Isolation Kit (Stemcell). Purity (>95%) of isolated B cells was verified by flow cytometry. For selected experiments, 1–2×105 B cells at baseline were stored in RNAprotect Cell Reagent (QIAGEN) and frozen at −80°C for subsequent RNA extraction.
B cell culture and TLR7/IFN stimulation
Human B cells freshly isolated from patients with SLE were resuspended at concentration 0.5×106 cells/mL in Iscove’s modified Dulbecco’s medium (IMDM) with 10% fetal calf serum (FCS), 1% antibiotic-antimycotic (Gibco) and 3 µM synthetic TLR7 agonist Resiquimod (R848, Invivogen) with/without 1000 IU IFNα (Enzo), together with iberdomide 1, 10 or 100 nM or vehicle control. At 5 days, cells were harvested for fluorescence-activated cell sorting (FACS) analysis, and supernatants stored for ELISA.
B cell culture and plasmablast differentiation
Human B cells freshly isolated from SLE patients were resuspended and cultured using a modified plasmablast differentiation protocol30 with HA-sCD40L (R&D) at 50 ng/mL cross-linked with 1 µg/mL anti-HA IgG (R&D), IL-2 at 20 U/mL, IL-10 at 50 ng/mL (Peprotech), IL-15 at 10 ng/mL (Peprotech) and TLR7 ligand R848 at 3 µM (Invivogen). Iberdomide (10 nM) or control vehicle was added at two timepoints, at either day 0 or 18 hours prior to harvest on day 4. After 5 days, cells were harvested for FACS, and supernatants stored for ELISA.
Flow cytometry and fluorescence activated cell sorting
Cultured B cells were characterised by flow cytometry on BD LSR Fortessa II with Live/Dead staining with Zombie NIR Dye (Biolegend), Fc blocking using Human Truestain FcX (Biolegend) and incubation with antibodies listed in online supplemental table 1). Samples undergoing plasmablast differentiation were cell sorted on BD FACSAria II into IgD+CD27- naïve B cells and CD20lowCD27+CD38+ plasmablasts. Immediately after sorting, cells were resuspended in RNAprotect Cell Reagent (QIAGEN) and frozen at −80°C for RNA extraction. Data were analysed using Flowjo software V10.
Supplemental material
ANA and immunoglobulin measurement
To measure antinuclear antibodies (ANA), 30 µL of neat supernatant was added to Hep-2 precoated microscope slides (BioDiagnostics). After washing in phosphate-buffered saline (PBS), bound immunoglobulins were detected with AF488 antihuman-IgG antibody (Invitrogen). Slides were read on an Olympus BX61 microscope interface with CellSense software. IgG and IgM were measured using total IgG and IgM ELISA Kits (Bethyl).
RNA sequencing
RNA was extracted using RNeasy Micro Kit (Qiagen) and concentration quantified on NanoDrop 2000c (ThermoFisher). RNA sequencing libraries were prepared using the NEBNext Ultra RNA Library Prep Kit for Illumina, validated on Agilent TapeStation and quantified using Qubit 2.0 Fluorometer (Invitrogen) and qPCR (KAPA). Libraries were sequenced on Illumina HiSeq 4000, using 2×150 bp paired end configuration, 50 million reads/sample.
RNA-seq data processing and analysis
Paired-end RNA-seq samples of 150 base pairs were trimmed to remove the Illumina adaptors using bbduk from BBMap package V.37.93. Transcripts were then quantified using Salmon version 0.13.1 with an index generated from Gencode release 29 transcriptome. Tximport version 1.13.10 was used to aggregate transcript level expression data to genes, and differential gene expression analysis was performed using DESeq2 V.1.25.9. P values were false discovery rate adjusted using Benjamini-Hochberg method. Genes regulated by Ikaros and Aiolos were identified via the Harmonizome website31 from ENCODE,32 CHEA Transcription Factor Targets,33 the Pathway Commons Protein-Protein Interactions (http://www.pathwaycommons.org/) and published literature5 14 34 (online supplemental table 2).
Supplemental material
Results
Iberdomide inhibits TLR7 and IFNα-induced activation of SLE B cells and abrogates autoantibody production
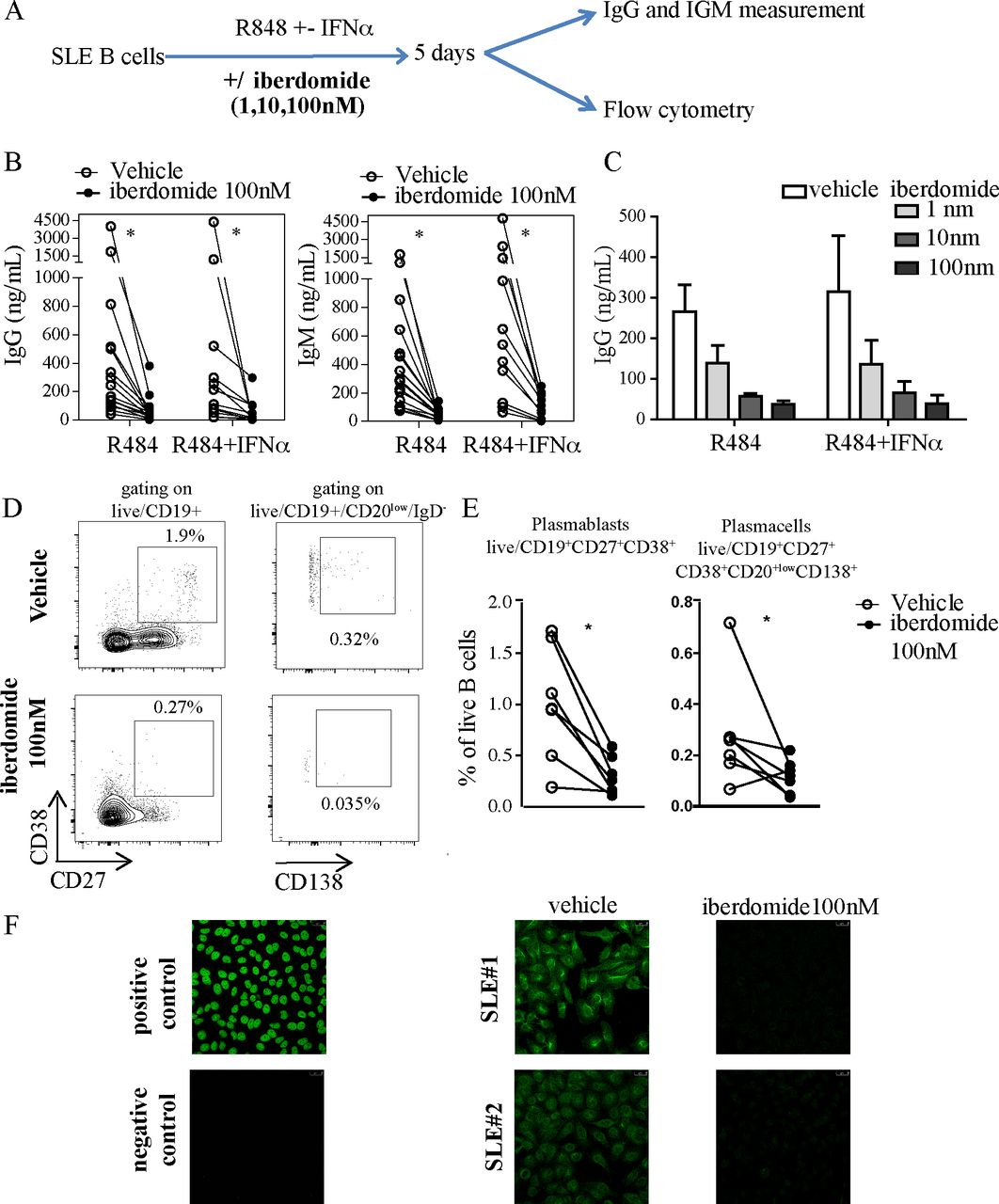
To explore the effects of therapeutic targeting of Ikaros and Aiolos on B cell activation in the context of SLE, we isolated B cells from blood samples from patients with SLE and stimulated them in vitro with TLR7 agonist resiquimod in combination with IFNα for 5 days, without or with the cereblon modulator iberdomide (figure 1A). TLR7 and IFNα stimulation of B cells promotes IgM+ response and expansion of autoreactive B cells35 and overactivity of both the TLR7 and IFNα system are established features of SLE.36–38 In our experiments, iberdomide significantly inhibited the production of IgG and IgM from B cells induced by resiquimod alone or resiquimod +IFNα (figure 1B), an effect that was shown to be dose-dependent, with both 10 and 100 nM inducing a significant reduction of Ig production compared with vehicle (figure 1C). Additionally, iberdomide induced a significant reduction in the percentage of CD19+CD27+CD38+ plasmablasts (iberdomide 0.3±0.18 vs vehicle 1.01±0.56, p=0.011) and CD20lowCD27+CD38+CD138+ plasma cells (iberdomide 0.12±0.06 vs vehicle 0.28±0.02, p=0.03) (figure 1D,E). Iberdomide was able to abrogate the production of NA) induced by resiquimod +IFNα, as measured by HEp2 immunofluorescence (figure 1F). Overall, these data show that iberdomide inhibits TLR7-mediated activation and differentiation of SLE B cells and abrogates the production of ANA induced in autoreactive B cells by TLR7 and IFNα triggering.
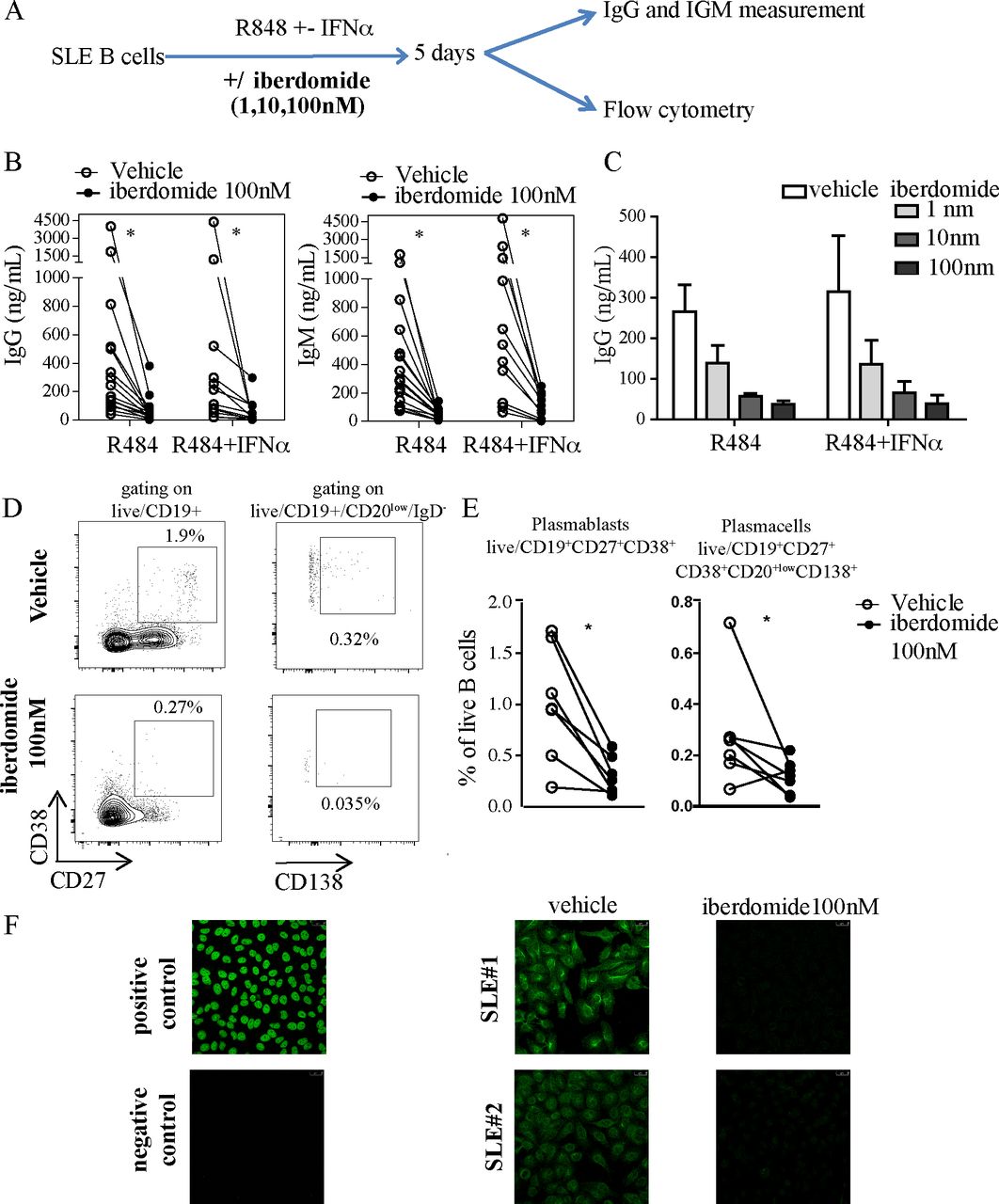
TLR7-induced and IFN-induced activation of SLE B cells. (A) Overview of experimental setup: B cells from SLE patients were triggered with TLR7 ligand R848 (Resiquimod) with iberdomide (1–10–100 nM) or vehicle as shown. After 5 days, cells were harvested and underwent flow-cytometry, while supernatants were used to measure ELISA. (B,C). IgG and IgM measured in the supernatants of cells stimulated as above with iberdomide at 100 nM (B) or 1–10–100 nM (C). (D) Representative dot-plots of flow cytometry after gating as shown and (E) cumulative results. (F) ANA measurement by IF in the supernatants of cells treated as in (A). n=16 patients with SLE in (B), 6 in (C), 7 in (E), with representative results in (D) and (F), p<0.05 Mann-Whitney in (B) and (E). ANA, antinuclear antibodies; SLE, systemic lupus erythematosus.
Iberdomide inhibits TLR7-mediated differentiation of plasmablasts from SLE B cells
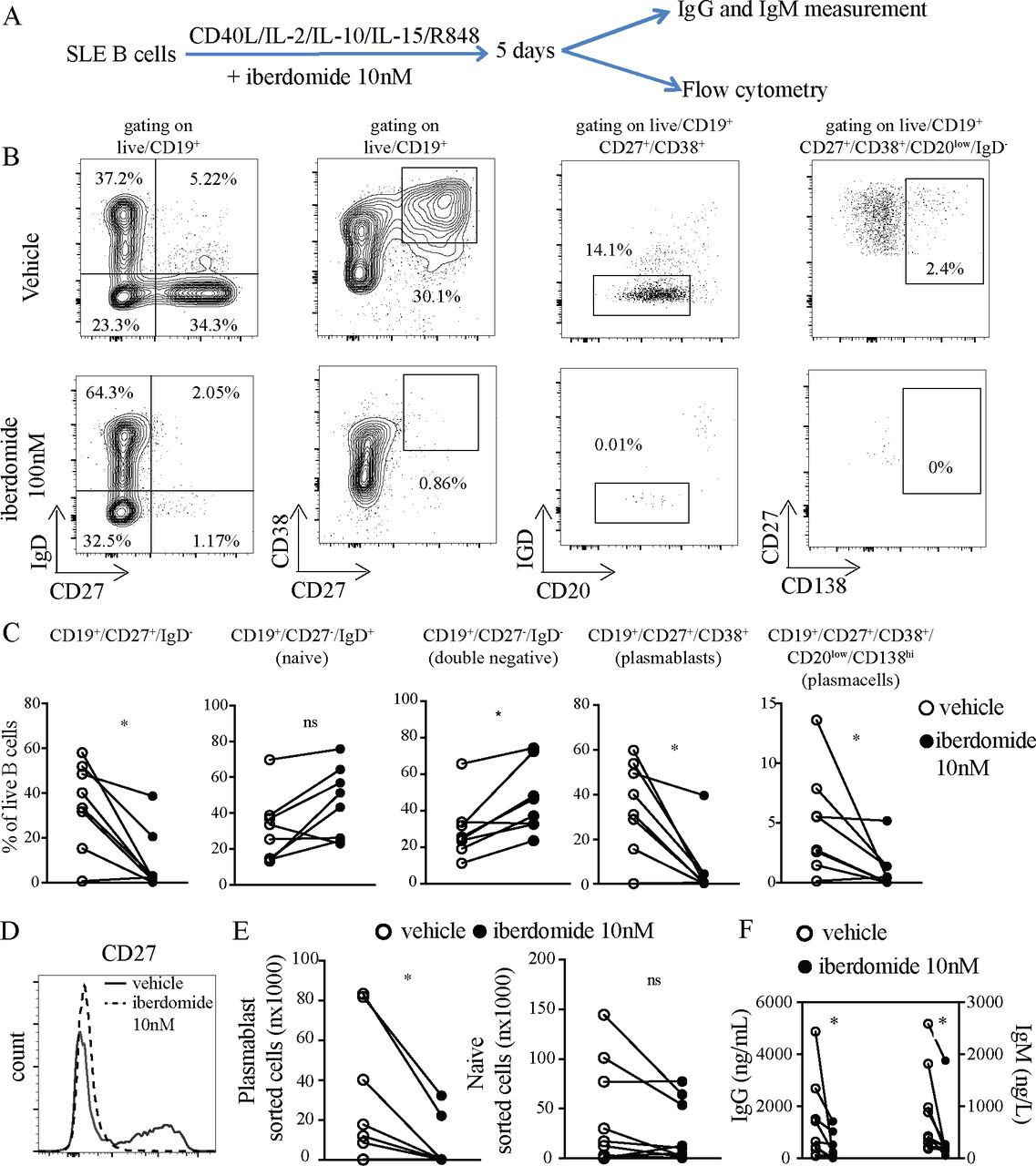
In order to explore the effects of iberdomide on the differentiation of B cells in SLE, we adapted a previously described method for differentiating B cells into plasma cells, which includes stimulation with IL2, IL10, IL15 and CD40L, substituting TLR7 agonist Resiquimod instead of TLR9 agonist CpG. Iberdomide at 10 nM or vehicle were added as indicated (the experimental setup is summarised in figure 2A). The dose of 10 nM was chosen since the dose-response experiments presented in figure 1C showed a significant functional effect at this dose, which is closer to pharmacologically efficacious levels in vivo. Treatment of B cells with iberdomide resulted in a significant reduction in the percentage of CD27+IgD- B cells (iberdomide 8.6±13.9 vs vehicle 34.9±19.3, p=0.028), without reducing the percentage of IgD+CD20low naïve B cells—if anything, treatment with iberdomide showed a trend towards an increase in the percentage of naïve B cells (iberdomide 45.6±19.9 vs vehicle 30.7±18.9, p=0.160) and led to a significant increase in double negative (DN) CD27-IgD- B cells considered part of the class-switched memory compartment (iberdomide 45.89±16.7 vs vehicle 29.4±16.2, p=0.015) (figure 2B,C). Additionally, iberdomide reduced the percentage of CD27+CD38+ plasmablasts (iberdomide 6.4±13.5 vs vehicle 34.9±20.1, p=0.013) and CD20lowCD27+CD38+CD138+ plasma cells (iberdomide 1.0±1.7 vs vehicle 4.0±4.3, p=0.010) (figure 2B,C). Accordingly, we observed profound suppression of CD27 expression, as shown in the representative dot-plots in figure 2B and the histogram in figure 2D. When cells underwent FACS, the absolute numbers of sorted plasmablasts was significantly lower in iberdomide-treated samples (iberdomide 6,085±12 215 vs vehicle 27,123±33 888, p=0.014), while the number of sorted naïve B cells was numerically lower, but not statistically significantly different (iberdomide 23 647±31 910 vs vehicle 44 849±50 678, p=0.09) (figure 2E). Furthermore, supernatants collected from B cells cultured in the presence of iberdomide had significantly lower levels of secreted IgG (iberdomide 272±327 vs 1357±1565, p=0.02) and IgM (iberdomide 204±72 vs vehicle 891±811, p=0.001) compared with vehicle (figure 2F).

Differentiation of SLE B cells into plasmablasts. (A) B cells from patients with SLE were triggered with HA-sCD40L (50 ng/mL) cross-linked with anti-HA IgG (1 µg/mL), IL-2 (20 U/mL), IL-10 (50 ng/mL), IL-15 (10 ng/mL) and TLR7 ligand R848 (3 uM), plus Iberdomide (10 nM) or control vehicle from day 0. (B) Representative dot-plots of flow cytometry of cells harvested after 5 days of culture. Gating on live CD19+. (C) Cumulative results of flow cytometry (D) representative histogram of CD27 expression by B cells treated with iberdomide or vehicle. (E) Absolute numbers of plasmablasts (CD27+CD38+) and naïve B cells sorted after 5 days of culture as above. (F) IgG and IgM measured in the supernatants of cells treated as above. n=8 patients with SLE in (C), (E) and (F), with representative results in (B) and (D). P<0.05 Mann-Whitney in C-E-F. SLE, systemic lupus erythematosus.
These results show that treatment with iberdomide significantly inhibits TLR7 mediated differentiation of SLE B cells into plasmablasts and plasma cells in vitro. As a consequence, antibody production is significantly reduced. Importantly, there were no significant differences in the percentage and absolute numbers of naïve B cells, suggesting a lack of impact on the overall survival of B cells, but rather a specific inhibition of their differentiation into plasmablasts.
Effects of Iberdomide on differentiated plasmablasts
To investigate the effects of iberdomide-mediated IKZF1 and IKZF3 inhibition on the transcriptional profiles of SLE plasmablasts, SLE B cells were differentiated into plasmablasts using TLR7, IL2, IL10, IL15 and CD40L costimulation as before. However, since addition of iberdomide throughout the culture completely blocked B cell differentiation into plasmablasts, in order to dissect its effects on transcriptional programmes separate from apoptosis programmes, in the next set of experiments iberdomide was only added 18 hours prior to cell sorting (at day 4) as shown in figure 3A. Iberdomide added to SLE B cells on day 4 did not impact the percentage of CD27+IgD- B cells, CD27+CD38+ plasmablasts, CD20lowCD27+CD38+CD138+ plasma cells and naïve B cells (figure 3B,C) nor the expression of CD27 on B cells (figure 3D). Accordingly, the absolute numbers of sorted plasmablasts and naïve B cells were unaffected (figure 3E). Iberdomide given briefly on day 4 did not alter production of IgG and IgM (figure 3F). Overall, this shows that treatment of in vitro differentiated plasmablasts with iberdomide for a shorter time did not affect the percentage and absolute numbers of B cell subsets, thus enabling transcriptomic analysis of iberdomide-treated plasmablasts to assess which genes are immediately impacted downstream of co-inhibition of IKZF1 and IKZF3.

Modulation of SLE B cell differentiation. (A) B cells from patients with SLE were triggered with HA-sCD40L (50 ng/mL) cross-linked with anti-HA IgG (1 µg/mL), IL-2 (20 U/mL), IL-10 (50 ng/mL), IL-15 (10 ng/mL) and TLR7 ligand R848 (3 uM) for 4 days, then Iberdomide (10 nM) or control vehicle was added at day 4 for 18 hours. (B) Representative dot-plots of flow cytometry experiments with cells harvested after 5 days of culture. Gating on live CD19+. (C) Cumulative results of flow cytometry (D) representative histogram of CD27 expression by B cells treated with iberdomide or vehicle. (E) Absolute numbers of plasmablasts (CD27+CD38+) and naïve B cells (IgD+CD20 low) sorted after 5 days of culture as above. (F) IgG and IgM measured in the supernatants of cells treated as above. n=16 patients with SLE in C-F-G, with representative results in (B) and (D). P<0.05 Mann-Whitney test in C-E-F. SLE, systemic lupus erythematosus.
Impact of Iberdomide on gene expression in naïve B cells and plasmablasts
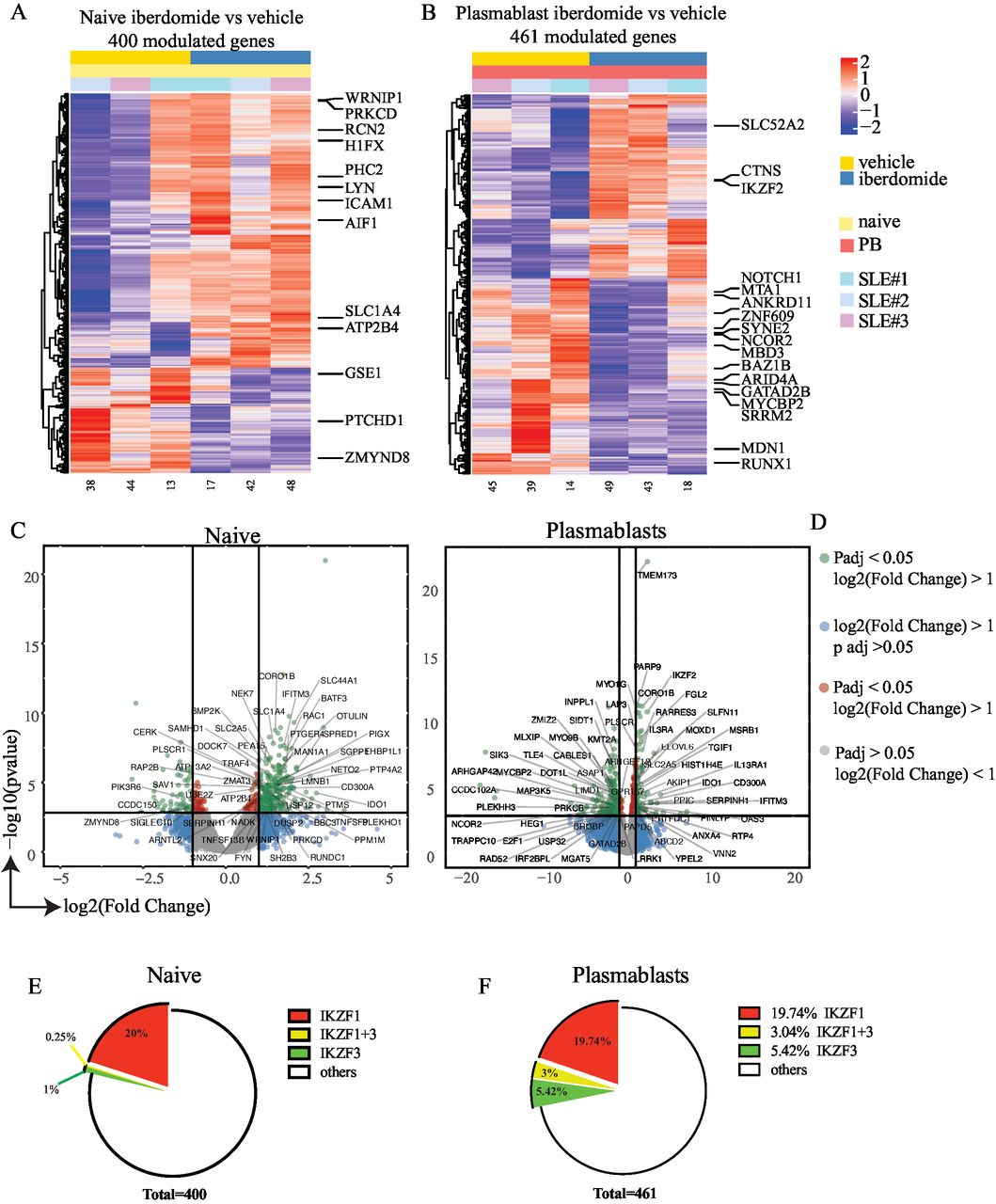
Sorted naïve B cells and plasmablasts treated with iberdomide were subjected to RNA-sequencing to determine the direct influence of iberdomide on gene expression. Initial comparison of differentiated plasmablasts and naïve B cells showed an upregulation of XBP1, IRF4, PRDM1 and the downregulation of BACH2, CIITA, PAX5 and BCL6, consistently with previously reported results in plasma cell differentiation,30 thus indicating that the modified differentiation protocol effectively activated plasmablast transcriptional programming (online supplemental figure 2). Then, we assessed cells treated with iberdomide, and we found a number of genes significantly modulated by iberdomide both in naïve B cells (400 genes) and plasmablasts (461 genes). Figure 4A,B show the results of unsupervised hierarchical clustering of the differentially expressed genes, with labelling for genes known to be regulated by Ikaros and Aiolos (online supplemental table 1), according to published transcription factor binding site profiling studies33 and literature on Ikaros and Aiolos.5 14 34 Genes which were significantly upregulated or downregulated by iberdomide in naïve B cells and plasmablasts are shown in the volcano plot in figure 4C,D, again identifying multiple genes which are known targets of Ikaros and Aiolos. Geneset enrichment analysis using the Enrichr R package showed that the differentially genes were significantly associated with CHIP-Seq datasets for IKZF1 (p=6.82×10-9) and IKZF3 (p=0.0007) target genes. Overall, more than 20% of the differentially modulated genes were identified as known targets of Ikaros and Aiolos (figure 4E,F). Many of these genes are also known to be involved in the pathogenesis of SLE (eg, NOTCH1, ZNF609, NCOR2, MYCBP2, SRRM2, MDN1, RUNX1, IGKV2D-10; IGLV3-17; IGHV1-45, CASP1, FGL2). Additional gene set enrichment analysis using EnrichR39 was performed in order to identify pathways modulated by iberdomide using the Reactome pathway database. Analysis of the most significant pathways downregulated by iberdomide in plasmablasts (online supplemental figure 1) identified pathways involved in chromatin modification and organisation as well as cell signalling (Rho GTPase, G-alpha), apoptosis (NRAGE signals death through c-Jun N-terminal kinase (JNK)), cytokine signalling (IL-2, IL-3) and cell cycle (BMAL1:CLOCK, p75 NTR, beta-catenin complex, VEGFR mediated cell proliferation). Overall, these data suggest that treatment of SLE B cells with iberdomide significantly affects gene expression downstream of Ikaros and Aiolos, thus modulating a number of critical cellular survival pathways in differentiating B cells.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Modulation of gene expression by iberdomide. (A,B) Hierarchical clustering of differentially modulated genes from RNA sequencing of sorted naïve B cells (A) and plasmablasts (B) after treatment as shown in figure 3(A). (C,D) Volcano plots showing genes that are significantly modulated by iberdomide in naïve B cells (C) and plasmablasts (D). Labelling is showing genes known to be modulated by Ikaros and Aiolos. (E,F) percentage of the total genes modulated in naïve B cells (E) and plasmablasts (F) that are known to be targets of Ikaros and Aiolos or both. n=3 patients with SLE from three independent experiments. SLE, systemic lupus erythematosus.
Discussion
In this study, we investigated the effects of pharmacologically targeting the transcription factors Ikaros and Aiolos using the cereblon modulator iberdomide on B cells isolated from patients with SLE. First, when B cells were triggered with the TLR7 ligand Resiquimod and IFNα, iberdomide could inhibit their activation and antibody production, including the production of autoantibodies (ANA). Additionally, iberdomide abrogated the in vitro differentiation of B cells into plasmablasts induced by IL-2/IL-10/IL-15/CD40L/Resiquimod, a protocol for the differentiation of plasmablasts that we adapted to include triggering with TLR7 agonist instead.30 Finally, when added to already differentiated plasmablasts, iberdomide induced a significant modulation of gene expression both in naive B cells and plasmablasts. Overall, these data support targeting of Ikaros and Aiolos by iberdomide as a therapeutic strategy in SLE, because of its ability to inhibit B cell activation and differentiation.
This is in line with previously published data, showing that (i) iberdomide treatment in vitro could reduce Ikaros and Aiolos expression in SLE B cells and inhibit the differentiation of B cells into plasmablasts induced by IL-2/IL-21/BAFF27 and (ii) the administration of iberdomide to healthy volunteers could reduce Ikaros and Aiolos levels in immune cells and inhibit anti-dsDNA and antiphospholipid antibody production from SLE mononuclear cells.28 Similar results have been shown for another thalidomide analogue, lenalidomide, which inhibited the generation of preplasmablasts and early plasma cells, moderately affected plasmablast production and inhibited long-lived plasma cell generation, but did not impair their long-term survival once generated.22
In our experiments, we further explored the effects of iberdomide on B cell activation by triggering B cells with the TLR7 ligand Resiquimod. TLR7 is highly relevant for the pathogenesis of SLE because of its ability to recognise RNA, released as a consequence of defective clearance of apoptotic debris, which represents a key pathogenetic mechanism in SLE40 and other autoimmune diseases.41 Overactivity of the TLR7 and IFNα system are well-established features of SLE.36–38 Additionally, while different triggers, including also CD40L/IL21, are able to support B cell survival and proliferation in culture, only TLR7 and IFNα have been shown to promote IgM+ response and expansion of autoreactive B cells and production of ANA in otherwise non-autoreactive healthy B cells, as TLR7 and IFNα stimulation of B cells.31 Except for a few genetic studies linking polymorphisms of IKZF1 and IKZF3 to SLE,42 the exact function of Ikaros family of transcription factors in the development of autoimmunity remains to be clarified. Only one recent study has investigated the association of Ikaros family of transcription factors with TLR signalling.15 The authors demonstrated hyperactive TLR signalling accompanied by the inability to induce B cell receptor anergy in Ikaros-deficient mice, thus leading to a predisposition to autoimmunity. Interestingly, animals with germinal centre B cell-specific Ikaros deletion developed neither autoimmunity nor germinal centre B cells, indicating that the loss of Ikaros stringently blocked germinal centre B cell development. This points towards a double-faceted function of Ikaros in the development of autoimmunity: on one hand, Ikaros can suppress B cell auto-reactivity, thus ‘protecting’ against autoimmunity; on the other hand, it is also essential for the activation and differentiation of B cells, as confirmed by the observation that Ikaros-deficient mice lack circulating plasma cells. Interestingly, Ikaros-deficient mice had signs of systemic autoimmunity but did not develop autoimmune disease, suggesting that additional factors are needed for the development of full-blown autoimmunity. Similarly, coding mutations of IKZF1 in humans mainly present with hypogammaglobulinaemia, while only a subset of patients develop autoimmunity.43–46 Overall, this suggests that Ikaros acts as a complex modulator of B cell activation and therefore represents a valuable target in autoimmune diseases. Accordingly, our experiments indicate that the addition of iberdomide throughout the whole duration of B cell culture was able to abrogate differentiation of B cells into plasmablasts and plasma cells, comparable to observations in Ikaros-deficient animals, which lack circulating plasma cells.
Importantly, iberdomide did not affect the overall survival of B cells, as the absolute number of naïve B cells and the percentage of naïve B cells did not change significantly and, if anything, higher levels of naïve B cells were found in iberdomide-treated patients, potentially a consequence of blockade of their differentiation. We observed a significant increase of the IgD-CD27- (DN) population on treatment with iberdomide. DN B cells,47 which are considered part of the memory compartment since they are class-switched, are expanded in SLE patients with high disease activity.48 Recent studies have shown that subclasses of DN B cells (DN2), which are TLR7 hyper-responsive, may promote SLE pathogenesis via their propensity to differentiate into plasmablasts following TLR7 activation.49 Overall, the relative increase in naïve B cells and DN B cells indicates that iberdomide is able to inhibit the TLR7 driven differentiation of B cells into plasmablasts.
These results consolidate previous data on B cells activated with CD40L and BAFF,27 providing further insights into the relevance of targeting Ikaros and Aiolos in patients with SLE because of the ability to inhibit TLR7 mediated B cell differentiation. The TLR7-driven differentiation protocol used in our study led to the expansion of both DN cells and plasmablasts in the absence of iberdomide. However, administration of iberdomide caused a drop in plasmablast number without affecting significantly naïve and DN B cells that were relatively increased. This observation supports the notion that iberdomide is not merely an anti-B cell compound, but through Ikaros and Aiolos downregulation, targets specifically plasmablast differentiation programming from naïve and DN cells, thus apparently halting the differentiation at that level. However, further studies are required to show whether iberdomide can block DN memory B cells expanded in SLE from differentiating into plasmablasts following TLR7 ligation. Although the lack of differences in the number of sorted naïve (IgD +CD27-) B cells clearly suggests that iberdomide does not impact B cell survival, a limitation of our study is that due to sample size and limited cell numbers we were not able to perform formal proliferation assays or use counting beads for additional accuracy.
Addition of iberdomide at a later stage to already differentiated plasma cells (18 hours before sorting), induced a modulation of gene expression both in naïve B cell and plasmablasts. This specific time-point was chosen based on previous time-course experiments, showing that treatment of B cells with iberdomide at 10 nM induced an inhibition of Ikaros and Aiolos lasting up to 24 hours.28 By performing RNA-sequencing of sorted naïve B cells and plasmablasts treated with iberdomide or control vehicle, we demonstrated that postactivation of the plasmablast transcriptional programme, iberdomide modulates a number of genes known to be regulated by Ikaros and Aiolos. Importantly, over 20% of genes that were modulated by iberdomide are known to be direct targets of Ikaros and Aiolos, in line with the known ability of iberdomide to induce the degradation of Ikaros and Aiolos.26 Although this is an exploratory analysis on a small number of patients, unsupervised hierarchical clustering identified a number of differentially expressed genes, many of which are known to be regulated by Ikaros/Aiolos, but also involved in the pathogenesis of SLE. For example, we identified: NOTCH1, whose blockade ameliorates SLE in animal models;50 ZNF609, a circular RNA implicated in lupus nephritis;51 NCOR2, relevant for the regulation of interferon production;52 MYCBP2, a newly described E3 ubiquitin-protein ligase;53 SRRM2, a splicing factor gene, with polymorphisms linked to SLE;54 MDN1, a human chaperone recently identified as a relevant factor in autoimmunity;55 RUNX1, a transcription factor associated with a number of autoimmune conditions, including rheumatoid arthritis, psoriasis and SLE.56
Similarly, analyses of differentially regulated genes in plasmablasts treated with iberdomide identified a number of genes known to be involved in the pathogenesis of SLE, such as immunoglobulin related genes (eg, IGKV2D-10; IGLV3-17; IGHV1-45); CASP1, encoding caspase-1, shown to be essential for the development of lupus in animal models;57 FGL2, fibrinogen-like 2, for which deletion leads to autoimmune glomerulonephritis.58
Additionally, pathway analysis identified a number of key cellular pathways that were downregulated by iberdomide in plasmablasts and may be important for plasmablast survival (inhibition of cell death signalling via JNK) during B cell differentiation and maturation and plasma cell survival in the bone marrow niche (beta catenin signalling). These data are consistent with previous known effects of targeting Ikaros and Aiolos leading to increased IL-2 signals. RNA-Seq analysis suggests that the inhibition of plasmablast/plasma cell differentiation and/or cell death of differentiating plasmablasts observed in our earlier experiments may be due to increased apoptosis in developing plasmablasts through increased death signals via JNK and alteration of cell cycle. Beta catenin is important for plasma cell survival via communication with bone marrow niche through syndecan-1 (also known as the plasma cell marker CD138),59 and so disruption of the beta catenin Wnt system may also lead to reduced plasma cell survival. Overall, these data indicate that iberdomide mediated degradation of Ikaros and Aiolos regulates key genes and pathways relevant for aberrant plasmablast differentiation during SLE pathogenesis.
These in vitro findings are also consistent with the recently reported clinical results for iberdomide in a SLE phase IIb study. Iberdomide showed significant efficacy in the treatment of active SLE, with enhanced effects observed in two biomarker-defined populations (Aiolos-High and Type 1 IFN-High), providing a rationale for this novel mechanism in SLE.60 In this trial, a significant decrease in anti-dsDNA antibodies was observed only after 24 weeks of iberdomide treatment.61 If there were a direct effect on plasmablast survival and immunoglobulin production, a more immediate decrease in anti-dsDNA antibodies would have been observed in this clinical trial. These clinical observations match the pattern of expression of cereblon, the molecular target of iberdomide, as it was found to be highly expressed in naïve B cells but downregulated once the differentiation in plasmablast is achieved.
In conclusion, we show that the cereblon modulator iberdomide inhibits the activation and differentiation of B cells and modulates the gene expression in differentiating plasmablasts from SLE patients, thus supporting the therapeutic targeting of Ikaros family transcription factors in SLE and potentially other autoimmune conditions.
Acknowledgments
We would like to thank all patients who were enrolled in the study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Twitter @FeliceRivellese
FR, SM-S and DM contributed equally.
Contributors Study conception and design: FR, DM, ML, PS, CP. Acquisition and analysis of experimental data: FR, SM-S and DM. FR wrote the first draft of the manuscript, DM and ML contributed to its revision. All authors contributed to the interpretation of the data, to the critical revision of the manuscript and approved its final version. ML had full access to all of the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Funding This investigator-led study was funded by an unrestricted grant from Celgene Corporation, now Bristol Myers Squibb.
Competing interests FR, MB, CP and ML received unrestricted grant support from Celgene. PS is an employee and shareholder of Celgene Corporation, now part of Bristol Myers Squibb.
Patient consent for publication Not required.
Ethics approval This study was conducted in accordance with the ethical principles of the Declaration of Helsinki and Good Clinical Practice guidelines and was approved by UK local ethical committee (REC reference 17/WS/0172).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available on reasonable request to the corresponding author. This includes detailed protocol for B cell stimulation and differentiation and experimental raw data (eg, RNA sequencing).
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.