Article Text

Abstract
Objectives The relationship of immune dysregulation and autoantibody production that may contribute to systemic lupus erythematosus (SLE) pathogenesis is unknown. This study evaluates the individual and combined contributions of autoantibodies, type I interferon (IFN-α) activity, and IFN-associated soluble mediators to disease development leading to SLE.
Methods Serial serum specimens from 55 individuals collected prior to SLE classification (average timespan=4.3 years) and unaffected healthy controls matched by age (±5 years), gender, race and time of sample procurement were obtained from the Department of Defense Serum Repository. Levels of serum IFN-α activity, IFN-associated mediators and autoantibodies were evaluated and temporal relationships assessed by growth curve modelling, path analysis, analysis of covariance and random forest models.
Results In cases, but not matched controls, autoantibody specificities and IFN-associated mediators accumulated over a period of years, plateauing near the time of disease classification (p<0.001). Autoantibody positivity coincided with or followed type II IFN dysregulation, preceding IFN-α activity in growth curve models, with elevated IFN-α activity and B-lymphocyte stimulator levels occurring shortly before SLE classification (p≤0.005). Cases were distinguished by multivariate random forest models incorporating IFN-γ, macrophage chemoattractant protein (MCP)-3, anti-chromatin and anti-spliceosome antibodies (accuracy 93% >4 years pre-classification; 97% within 2 years of SLE classification).
Conclusions Years before SLE classification, enhancement of the type II IFN pathway allows for accumulation of autoantibodies and subsequent elevations in IFN-α activity immediately preceding SLE classification. Perturbations in select immunological processes may help identify at-risk individuals for further clinical evaluation or participation in prospective intervention trials.
- Systemic Lupus Erythematosus
- Autoantibodies
- Autoimmunity
- Chemokines
- Cytokines
Statistics from Altmetric.com
Introduction
Systemic lupus erythematosus (SLE) is marked by pathogenic autoantibody production and immune dysregulation, resulting in organ damage. Patients may accrue organ damage by the time they reach disease classification,1 making early intervention an attractive approach to curtailing excessive morbidity. Antinuclear autoantibodies (ANAs) might help identify future patients since SLE-associated autoantibodies accumulate prior to clinical disease.2 ,3 However, up to 14% of healthy individuals, particularly women aged 40–49 years, are ANA positive yet never develop lupus or another autoimmune rheumatic disorder.4 Therefore, ANA positivity is likely accompanied by other immunological changes contributing to pathogenesis and may help distinguish patients at risk of disease transition.
Elevated type I interferon (IFN-α) activity is associated with SLE pathogenesis,5 including DNA-protein and RNA-protein binding autoantibody specificities6 and B-lymphocyte stimulator (BLyS) induction.7 BLyS, a key regulator of B-cell survival and differentiation, contributes to autoantibody production and class switching.8 Both IFN-α9 and BLyS8 have been recent lupus therapeutic targets. Although BLyS blockade may reduce disease activity in some lupus patients,8 clinical trials using IFN-α blocking agents have had modest clinical outcomes to date, despite evidence of decreasing IFN-α gene signatures.9
In addition to type I IFN, multiple genes that contribute to activation of type II IFN (IFN-γ) pathways are associated with SLE.10 Furthermore, IFN-γ can drive both type I IFN11 and BLyS production.12 Bridging innate and adaptive immunity, IFN-γ is expressed by multiple innate cell types13 and primes macrophages and dendritic cells to respond to toll-like receptor (TLR) signals.11 IFN-γ perpetuates Th1-type adaptive cellular responses, recruiting cells to sites of inflammation by stimulating the secretion of such chemokines as IFN-gamma-inducible protein-10 (IP-10) (CXCL10), monokine induced by IFN-gamma (MIG) (CXCL9) and macrophage chemoattractant protein (MCP)-3 (CCL7).11 Treating patients with SLE with the anti-IFN-γ monoclonal antibody AMG 811 reduces IP-10 levels,14 which itself has become a potential rheumatic disease therapeutic target.15
Together, these observations suggest that dysregulation of type I/type II IFNs and autoantibody accumulation may contribute to SLE pathogenesis. However, levels of IFN-associated mediators in preclinical SLE and possible interactions between IFN-α activity, type II IFN pathways and autoantibody accumulation in the transition to clinical disease are not understood. We sought to address this knowledge gap by analysing longitudinal serum samples collected prior to and concurrent with SLE classification. In this first preclinical longitudinal study to assess the temporal relationship between IFN-associated soluble mediators, the accrual of lupus-associated autoantibodies and increased type I IFN activity, our findings suggest that perturbations in combined immunological processes precede clinical classification and can help identify individuals at risk of lupus development.
Methods
Study population and serum samples
A subset of individuals in the Department of Defense Serum Repository (DODSR, n=55) was selected for this study based on availability of serum samples collected prior to and at/after disease classification. Demographic and clinical information was collected at the time of sample procurement. Two-thirds of subjects were female, 55% were African–American (AA) (25% European–American) and mean (SD) age at SLE classification was 29.4 (6.0) years.
Cases were matched to healthy controls by age (±5 years), gender, race and time of sample procurement. Demographics are shown in online supplementary table S1.
Serum autoantibody detection, IFN-α activity measurement and soluble mediator detection
Autoantibodies to dsDNA, chromatin, Ro/Sjögren's-syndrome-related antigen A (SSA), La/SSB, Sm, SmRNP complex and ribonucleoprotein (RNP) were detected using the xMAP BioPlex 2200 multiplex system (Bio-Rad Technologies, Hercules, California, USA).16 WISH Reporter cells (CAT# CCL125; ATCC) were used to measure the ability of sera to induce type I IFN-related gene transcription.6 Serum BLyS levels were determined by ELISA (R&D Systems, Minneapolis, Minnesota, USA) and IP-10, IFN-γ, MCP-3, MIP-1α, and MIG assessed by xMAP multiplex assays (Affymetrix-eBioscience, Santa Clara, California, USA)17 (see online supplementary methods).
Statistical analyses
Associations between time to SLE classification versus IFN-α activity, autoantibodies and IFN-associated mediators were analysed by Kruskal–Wallis testing with Dunn's multiple comparison. Differences between cases and controls with respect to accumulation of autoantibody specificities and IFN-associated mediators were determined by Wilcoxon matched-pairs test. Within-individual correlations between autoantibody specificities or IFN-associated mediator levels and IFN-α activity were computed by comparing the residual variance from an unconditional means model to that of a Gaussian mixed model, where the variable of interest was included as a predictor. The resulting pseudo R2 value is defined as (var(unconditional)-var(growth))/var(unconditional). Growth curve modelling, path analysis, analysis of covariance (ANCOVA) and random forest (RF) predictive modelling assessments were performed (see online supplementary methods). Analyses were performed using GraphPad Prism 6.02 for Windows (GraphPad Software, San Diego, California, USA), SAS STAT V.9.3 (SAS Institute, Cary, North Carolina, USA) and R 2.15.3 (R Foundation for Statistical Computing, Vienna, Austria).
Results
IFN-α activity increases before SLE disease classification
IFN-α is genetically and serologically associated with SLE pathogenesis.5 ,6 ,18 We evaluated longitudinal changes in serum IFN-α activity over three longitudinal serum samples procured from 55 cases in the DODSR, covering symptom-free, pre-classification, and at/after SLE classification periods. IFN-α activity increased as individuals approached disease classification (p<0.001 median difference=2.32, 95% CI 0.51 to 2.94, figure 1A). Only 20% of cases had increased serum IFN-α activity ≥4 years before SLE classification, while 44% had elevated serum IFN-α activity ≤2 years before classification, and 75% had increased serum IFN-α activity at classification.

Interferon (IFN)-α reporter activity levels increase as individuals move towards systemic lupus erythematosus (SLE) classification. Increases in serum IFN-α activity score by time before SLE classification (A) and by autoantibody status (B) are shown (mean±SEM). IFN-α activity score ≥1 is considered positive (dashed line). Per cent of case samples positive for IFN-α activity is listed below each time point. *p<0.05, ***p<0.001, Kruskal–Wallis with Dunn's multiple comparison. Demographics (C) and number of positive autoantibodies (D, mean±SEM; *p<0.05, unpaired t test) in SLE cases that are ever IFN-α activity positive versus SLE cases that remain IFN-α activity negative are also shown. Other=Asian/Pacific Islander, American Indian/Alaskan Native or multiracial.
Since patients approaching SLE classification accumulate DNA-associated and RNA-associated autoantibodies,3 we examined the relationship between serum IFN-α activity and autoantibody positivity (figure 1B). Compared with the autoantibody negative time point, serum IFN-α activity was increased with autoantibody positivity (p=0.055 median difference=0.58, 95% CI 0.0 to 1.64) and further increased at/after disease classification (p<0.001 median difference=2.34, 95% CI 0.65 to 3.26). Of the 55 cases, 41 (75%) had at least one sample with elevated IFN-α activity, with no significant differences in gender, ethnicity or SLE classification age (figure 1C). Cases with positive serum IFN-α activity had more DNA-associated and RNA-associated autoantibody specificities (3.8±2.1) than cases without elevated IFN-α activity (2.4±2.6) (p=0.050; figure 1D), and the number of autoantibody specificities correlated with serum IFN-α activity level (Spearman r=0.462, p<0.001). These results imply a biological relationship between IFN-α activity and autoantibody accrual during preclinical SLE.
Elevation of IFN-α activity follows autoantibody accumulation
The number of autoantibody specificities increased preclinically and plateaued near disease classification in individuals who developed classified SLE, but not matched healthy controls, (p<0.001; figure 2A). The levels of each autoantibody specificity showed significant within-individual correlations with IFN-α activity (figure 2B). We evaluated the temporal relationship between IFN-α activity and DNA-associated and RNA-associated autoantibody production using growth curve linear mixed models, which account for the contribution of the longitudinal samples acquired at varying times relative to disease classification. All RNA-associated (figure 2C) and DNA-associated (figure 2D) autoantibodies, except anti-dsDNA, appeared before elevation of IFN-α activity and SLE classification in all cases (see online supplementary figure S1); no differences in the frequency of autoantibody specificities and IFN-α activity were noted between AA and non-AA cases (see online supplementary table S2). Anti-dsDNA antibodies typically coincided with elevated IFN-α activity (mean appearance at −2.40 vs −2.49 years), although only 37 (67%) of cases were dually positive for serum IFN-α activity and anti-dsDNA. Therefore, autoantibody positivity and accumulation of lupus-associated autoantibody specificities can be detected prior to increases in IFN-α activity.
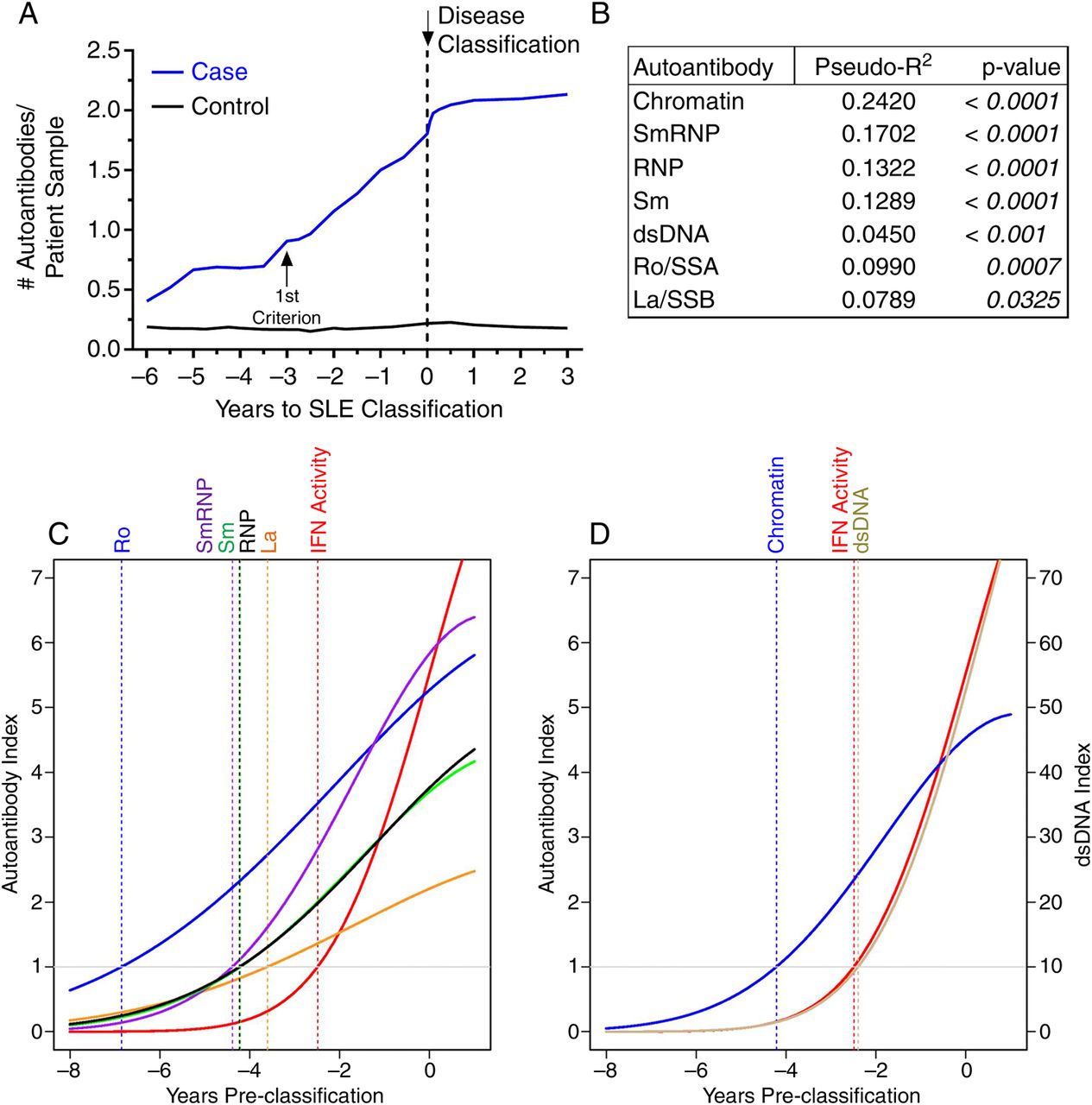
Most autoantibodies are detected prior to increases in interferon (IFN)-α activity in patients with systemic lupus erythematosus (SLE). (A) Increase in the number of serum autoantibodies by time before SLE in 55 cases compared to matched healthy controls, p=0.0002 by Wilcoxon matched-pairs test. Time of SLE classification (dotted line, down arrow) is shown. Within-individual correlations of IFN-α activity versus level of autoantibody positivity are presented, as described in the ‘Methods’ (B). Temporal relationships, using growth models, of IFN-α activity with Ro/SSA, La/SSB, Sm, SmRNP and ribonucleoprotein (RNP) (C) and with double-stranded DNA (dsDNA) and chromatin autoantibodies (D) are shown. The positive cut-off values are dsDNA >10 IU/mL and >1 autoantibody index for all other specificities by xMAP multiplex assays. Time of positivity (years pre-classification): dsDNA (−2.40), IFN-α activity (−2.49), La/SSB (−3.61), Sm (−4.21), chromatin (−4.21), RNP (−4.23), SmRNP (−4.39), and Ro/SSA (−6.85).
IFN-associated soluble mediators correlate with IFN-α activity and accumulation of autoantibodies
Increasing serum IFN-α activity observed in patients approaching SLE classification (figure 1A,B) may reflect inflammatory processes that stimulate IFN target gene expression.19 We evaluated serum levels of IFN-associated mediators in pre-classification cases compared with time-of-procurement matched controls. Significant within-individual correlations existed between IFN-α activity and levels of IFN-γ (p<0.001, pseudo R2=0.151), IP-10 (p<0.001, pseudo R2=0.243), BLyS (p<0.001, pseudo R2=0.322) and MCP-3 (p=0.002, pseudo R2=0.150), with a moderate correlation between IFN-α activity and MIG levels (p=0.040, pseudo R2=0.096; see online supplementary figure S2A). Further, serum IFN-γ (p<0.001 median difference=−7.65, 95% CI −8.04 to −6.45), IP-10 (p=0.002 median difference=−26.45, 95% CI −41.95 to −9.96) and MCP-3 (p<0.001 median difference=−344.3, 95% CI −351.5 to −192.9) concentrations were significantly higher in cases ≥4 years pre-classification compared with matched controls, with additional increased levels as patients moved towards classification (p≤0.001; see online supplementary figure S2B–E). Control samples maintained consistent levels of the IFN-associated soluble mediators, whether assessed with respect to time of SLE classification in matched cases, relative time of sample procurement or order of sample procurement (see online supplementary figure S3).
Given that type II IFN-associated mediators were elevated several years prior to SLE classification, we hypothesised that they may increase prior to or concurrent with the detection of SLE-associated autoantibodies. Elevated IFN-associated soluble mediators in each longitudinal sample were defined using cut-offs established by receiver operating characteristic curves that optimally differentiated cases versus controls (see online supplementary figure S4). The number of elevated IFN-associated inflammatory soluble mediators increased prior to and plateaued near disease classification in cases, but not matched controls (p<0.001 median difference=−0.81, 95% CI −1.88 to −0.49; figure 3A). Cases had a mean of 4.7 elevated mediators at the time of classification compared with 1.8 in controls. IFN-α activity and levels of IP-10, IFN-γ, BLyS and MIG showed significant within-individual correlations with the number of positive autoantibody specificities (p≤0.001; figure 3B), with an increased number of elevated IFN-associated mediators as cases moved from autoantibody-negative status (2.4 elevated mediators) to disease classification (4.7 elevated mediators; p≤0.001 median difference=−2.41, 95% CI −2.53 to −1.62). Importantly, levels of IFN-γ (p<0.001 median difference=−7.49, 95% CI −9.03 to −5.08, figure 3C) and IP-10 (p=0.007 median difference=−29.46, 95% CI −74.49 to −5.76, figure 3D) were significantly higher in autoantibody-negative cases compared with controls. However, levels of BLyS and MIG were not significantly elevated in cases until at/near the time of SLE classification (figure 3E,F). Because case samples were procured concurrently with matched control samples, longitudinal changes in serum IFN-associated soluble mediators in cases were not a consequence of time, but instead were likely related to the preclinical mechanisms that precipitated SLE disease classification (figure 3 and online supplementary figures S2 and S3). Together, these results provide further support that, like IFN-α activity, IFN-associated soluble mediators are dysregulated prior to clinical SLE. In addition, they suggest that IFN-γ and IP-10 increase prior to SLE-associated autoantibodies.

Interferon (IFN)-associated soluble mediators increase as individuals move towards systemic lupus erythematosus (SLE) classification and correlate with number of autoantibodies. Increase in number of serum IFN-associated mediators by time before SLE classification in 55 cases versus matched healthy controls, p=0.0002 by Wilcoxon matched-pairs test (A). Time of SLE classification (dotted line, down arrow) is shown. Within-individual correlations of IFN-associated serum soluble mediators versus number of positive serum DNA-associated and RNA-associated autoantibodies are shown, as described in ‘Methods’ (B). Serum soluble mediators IFN-γ (C), IP-10 (D), B-lymphocyte stimulator (BLyS) (E) and MIG (F) in cases by autoantibody status and in matched controls (Ctl) are shown (mean±SEM). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 by Kruskal–Wallis test with Dunn's multiple comparison (Cases) or by Mann–Whitney (Cases vs Ctl).
Type II IFN mediators and autoantibodies precede elevated IFN-α activity and SLE classification
The above findings, largely from univariate analyses of aggregate samples, suggest the following timing of events prior to SLE classification: dysregulation of select type II IFN mediators, followed by the detection and accrual of autoantibodies, and finally, the elevation of serum IFN-α activity. To evaluate this temporal relationship directly, on a per-patient basis, we performed path analysis using data from three longitudinal samples per individual to determine the relative timing of increased levels of IFN-associated soluble mediators, autoantibody positivity and elevated IFN-α activity.
We considered three ordinal relationships between autoantibody positivity and IFN-α activity: IFN-α activity before autoantibody positivity, autoantibody positivity before IFN-α activity and IFN-α activity concurrent with autoantibody positivity (figure 4A). Each model was ranked based on three fit indices. In the Akaike's information criterion (AIC) and root mean square error of approximation (RMSEA) goodness-of-fit indices, lower values (AIC, no fixed range; RMSEA, range 0–1) indicate a better model fit.20 In the lack-of-fit χ2 index, larger χ2 values (smaller p values) indicate a less parsimonious model and/or poorer fit.21 We found that autoantibody positivity preceding IFN-α activity was the top ranking model by all fit indices for each autoantibody specificity except anti-dsDNA (figure 4B, highlighted), consistent with the growth curve models (figure 2C,D). Anti-dsDNA positivity preceded IFN-α activity per the AIC fit index and coincided with IFN-α activity per RMSEA and χ2 model fit indices (figure 4B), consistent with the growth curve models showing anti-dsDNA positivity coinciding with increased IFN-α activity (figure 2D). Moreover, in 84% of individuals positive for both IFN-α activity and autoantibodies, at least one autoantibody specificity appeared prior to or concurrent with elevation of IFN-α activity (see online supplementary table S3).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Interferon (IFN)-associated mediators are elevated prior to or concurrent with autoantibody positivity, which precedes increases in IFN-α activity in patients with systemic lupus erythematosus (SLE). (A) Abbreviated path analysis diagrams showing posited temporal relationships tested in three longitudinal samples/patient (sample 1, sample 2, sample 3): positive IFN-α activity preceding autoantibody (AutoAb) detection, AutoAb detection preceding positive IFN-α activity or the simultaneous detection of both AutoAb and IFN-α activity positivity. Results comparing the timing of positive IFN-α activity versus the detection of lupus-associated antibody specificities are shown (B). (C) Abbreviated path analysis diagrams showing posited temporal relationships tested in three longitudinal samples/patient (sample 1, sample 2, sample 3): positive soluble mediator preceding AutoAb detection or IFN-α activity, AutoAb detection or IFN-α activity preceding soluble mediator or the simultaneous detection of both soluble mediator and AutoAb or IFN-α activity. Results comparing the timing of positive soluble mediator versus the detection of anti-Ro/SSA and IFN-α activity are shown (D). The highest model rank (highlighted) was determined by the lowest values of the goodness-of-fit model indices Akaike’s information criterion (AIC) and root mean square error of approximation (RMSEA) and the highest p value for the lack-of-fit (LOF) χ2 index. dsDNA, double-stranded DNA.
Next, we considered path analysis models similarly positing that IFN-associated soluble mediators preceded, followed or appeared concurrently with autoantibody positivity (specifically anti-Ro/SSA, the first autoantibody to be detected, figures 2C and 4B) and elevated IFN-α activity (figure 4C). We found that type II IFN mediators, IFN-γ and IP-10, were more likely to be elevated concurrent with or prior to anti-Ro/SSA positivity, respectively, and that both were elevated prior to the increase in IFN-α activity. In contrast, BLyS was more likely to be elevated after both anti-Ro/SSA positivity and elevated IFN-α activity (figure 4D). These findings corroborate the univariate analyses showing elevated IFN-γ and IP-10 in preclinical cases prior to autoantibody positivity, and continued increases of these mediators (with later BLyS elevation) coinciding with autoantibody accrual and SLE classification (figure 3 and online supplementary figure S2). Further supporting these results, most individuals were positive for IFN-γ (98%) and IP-10 (87%), along with MCP-3 (92%), prior to or concurrent with autoantibody positivity, while BLyS positivity occurred concurrent with or after autoantibody detection and serum IFN-α activity (87%; see online supplementary table S4).
Type II IFN mediators and autoantibodies predict elevated IFN-α activity and SLE classification
Because elevated serum IFN-α activity occurs proximal to the time of SLE classification, we applied ANCOVA analyses to determine the best predictors of IFN-α activity elevation. We evaluated whether IFN-associated soluble mediators, autoantibody specificities, gender and/or ethnicity might predict increased serum IFN-α activity and SLE classification. We found that the number of autoantibody specificities (p<0.001), female gender (p=0.010) and number of positive IFN-associated mediators (p=0.038) were significant predictors of increased IFN-α activity (table 1).
Gender, number of positive autoantibodies and number of interferon (IFN)-associated soluble mediators predict increase in IFN-α activity
Since several IFN-associated mediators were found to be elevated prior to or concurrent with the accumulation of autoantibody specificities and serum IFN-α activity before SLE classification, we sought to determine which preclinical factors best predicted SLE development. We used RF modelling22 to evaluate the following variables as possible predictors of future SLE classification: the presence of SLE-associated autoantibodies, elevated serum IFN-α activity and elevated levels of IFN-associated mediators. Interestingly, IFN-α activity was eliminated as a predictor variable because it did not make a significant, independent contribution to the prediction of SLE classification. Future SLE classification was best discerned by the concentrations of IFN-γ, MCP-3, anti-chromatin and anti-spliceosomal autoantibodies, even >4 years before classification (93% accuracy, out-of-bag (OOB) error=0.07; table 2). Thus, a combination of type II IFN mediators and autoantibodies accurately predicts SLE classification years in advance. Further, these findings suggest that early dysregulation of type II IFN mediators interacts with accumulation of autoantibody specificities, leading to elevated serum IFN-α activity and ultimately resulting in clinical disease and SLE classification.
Interferon (IFN)-associated mediators and select autoantibody specificities predict transition to systemic lupus erythematosus
Discussion
Understanding early lupus pathogenesis is critical for assessing and minimising the risk of transitioning to SLE. SLE is fundamentally a disease of immune dysregulation. Using a unique resource of well-characterised, longitudinal serum samples collected prior to and at/after SLE classification, we found that, like autoantibody positivity, IFN-α activity and elevated IFN-associated soluble mediators arise months or years before disease classification.
Type I IFN (IFN-α)5 and BLyS8 have been linked to SLE pathogenesis; our data show increased IFN-α activity and BLyS levels shortly before SLE classification, which may denote a turning point in pathogenesis, where positive feed-forward mechanisms amplify immune dysregulation past the threshold for inevitable transition to classified SLE. Elevated IFN-α activity and BLyS levels may arise from early dysregulation of type II IFN mediators interacting with the accumulation of lupus-associated autoantibody specificities. This temporal relationship is supported across multiple multivariate models, including growth curve models, path analysis and ANCOVA, placing autoantibody positivity before or near elevated IFN-α activity. Indeed, in our investigation, the number of positive autoantibodies was the most significant (p<0.001) predictor of increased IFN-α activity in cases. A recent study demonstrated IFN-α activity associated with autoantibody positivity, a small study (n=24) showed exacerbated type I IFN signatures in incomplete lupus erythematosus patients whose autoantibody specificities had already class-switched to the pathogenic IgG isotype23 and a number of studies have noted the ability of autoantibody-containing immune complexes to drive type I IFN activation.24–27 While IFN-α activity is clearly important in the transition to classified SLE, it may not be a leading factor for the initial development of lupus-associated autoantibody specificities. Indeed, roughly 25% of the patients in the current study did not have positive IFN-α activity, further highlighting the heterogeneous nature of SLE.
Preclinical detection of autoantibodies prior to increases in IFN-α activity and the modest clinical benefit from IFN-α blockade in recent clinical trials9 infer that other forms of immune dysregulation accompany IFN-α activity to allow for clinical disease. Previous evidence in lupus-like animal models and patients with SLE suggests that amplified crosstalk between innate cells and lymphocytes leads to breaks in tolerance, enabling the activation and persistence of autoreactive B cells.28 IFN-γ, a key mediator of such crosstalk, facilitates autoantibody production by modulating TLR regulation, antigen presentation and lymphocyte recruitment to germinal centres,29 where IFN-γ promotes IgG class switching and pathogenic autoantibody production.30 Our temporal analyses show elevated levels of IFN-γ and the chemokines IP-10 and MCP-3 in patients with preclinical SLE prior to IFN-α activity and to positivity for most autoantibodies. Moreover, IFN-γ and MCP-3 consistently determine transition to classified SLE, even >4 years before disease classification, supporting an early role for these IFN-associated mediators in SLE pathogenesis. In longitudinal studies of patients with undifferentiated connective tissue disease, increased levels of IFN-γ signal the transition from benign to pathological disease,31 and those who transition to SLE accumulate lupus-associated autoantibodies.32 Finally, IFN-γ can drive the production of IFN-α11 and BLyS,12 thereby promoting inflammation and further enhancing B-cell activation and (auto)antibody accumulation.30 ,33 Serum BLyS levels were not significantly elevated in cases compared with matched controls until shortly before SLE classification and after increases in serum IFN-γ, autoantibody positivity, and serum IFN-α activity.
This study provides critical new information to help identify ANA-positive individuals at the highest risk of transition to SLE. Through the measurement of systemic soluble mediators indicative of ongoing inflammation when detected in the periphery, including IFN-γ, IP-10, MCP-3 and BLyS, in addition to the detection of DNA-protein and RNA-protein binding autoantibodies, we can identify individuals who may need rheumatology referral or closer monitoring, findings further supported by similar perturbations in autoantibodies and cytokines in preclinical rheumatoid arthritis.34 ,35 In addition, results from this study will help identify individuals at significantly increased risk of SLE development with a high level of specificity, sensitivity and predictive accuracy, thereby providing critical information needed to design and implement prevention trials. Such prospective clinical trials would allow for parallel procurement of detailed clinical assessment and biological specimens to address a current limitation of this study. By understanding that IFN-associated pathway dysregulation occurs years prior to clinical symptoms with further increases closer to the time of clinical disease onset, we may be able to select and test potential early therapeutic agents, such as hydroxychloroquine,36–38 for individuals before irreversible organ damage has occurred. This may be potentially beneficial for those patients in whom organ damage occurs by the time they reach disease classification as early damage has been shown to be predictive of further damage, comorbidities and early mortality.39–41
Acknowledgments
The authors thank Krista Bean, Jourdan Anderson, Tim Gross and Wade DeJager for technical assistance, as well as Rebecka Bourn, PhD, and Angela, Andersen, PhD, for editorial assistance.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Handling editor Tore K Kvien
Contributors MEM, TBN, GCT, MPK, JBH and JAJ designed the study. MEM, JRA, TG, JMR, BFB, TBN and JAJ participated in data acquisition. MEM, RL, JRA, DAF, JMR, YDZ, TBN, JBH and JAJ participated in data analysis. All authors assisted with the development of the manuscript and gave final approval for publication. MEM, RL, JMR and JAJ had full access to data for the study. JAJ had the final responsibility for the decision to submit for publication.
Funding Research reported in this publication was supported by the National Institute of Allergy, Immunology and Infectious Diseases, Office of Research on Women's Health, National Institute of General Medical Sciences and the National Institute of Arthritis, Musculoskeletal and Skin Diseases under award numbers U01AI101934, U19AI082714, U54GM104938, P30GM103510, P30AR053483 and S10RR026735. This material is also the result of work supported with resources and the use of facilities through the Department of Veterans Affairs. Additional support was provided by the National Institute of Allergy, Immunology and Infectious Diseases and National Institute of Arthritis, Musculoskeletal and Skin Diseases under award numbers AI071651 and AR060861 (TBN). JBH would like to acknowledge support from the US Department of Veterans Affairs and NIH grants U01HG006828, UL1TR000077, R37AI024717, R21AI103980, P01AI083194 and P01AI049084.
Disclaimer This publication is the sole responsibility of the authors and does not represent the views of the National Institutes of Health or the Department of Veterans Affairs. The views expressed in this article are those of the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of the Army, US Armed Forces Department of Defense, or the US Government.
Competing interests None declared.
Ethics approval Experiments were performed in accordance with the Helsinki Declaration and approved by the Institutional Review Boards of the Oklahoma Medical Research Foundation and the Walter Reed National Military Medical Center.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement The data resulting from this study are available upon written request submitted to the corresponding author, subject to (1) the Material Transfer Agreement and Materials and Data Usage Agreement of the Oklahoma Medical Research Foundation, (2) the approval of the Armed Forces Health Surveillance Center and (3) documentation of all appropriate research training, including but not limited to human subjects research training.