Article Text

Abstract
Objectives To determine whether type 1 interferon (IFN) proteins in blood are associated with downstream type 1 IFN-inducible gene expression in blood from patients with myositis.
Methods IFNα, IFNβ and IFNω concentrations were measured by ELISA in 129 blood samples (from 93 patients with dermatomyositis (DM), inclusion body myositis, polymyositis and other muscle diseases and from 36 healthy volunteers). Their concentrations were correlated with their ability to stimulate type 1 IFN-inducible gene transcription in a functional assay for 123 of these samples and the type 1 IFN-inducible blood gene expression from 70 of the same samples.
Results Blood IFNβ concentration was uniquely associated with DM (p=0.0004), detectable in 64% of samples from patients with untreated or minimally treated DM and 35% of all DM samples compared with 6% of other inflammatory myopathy and 6% of healthy volunteer samples. Blood IFNβ, but not IFNα or IFNω, correlated with high blood type 1 IFN-inducible gene expression (p=0.01). Healthy volunteer samples with a high ELISA signal for IFNα and IFNω lacked functional bioassay activity and such a signal was confirmed as artefactual.
Conclusion Elevated blood IFNβ protein concentration is associated with DM. Systemic and local production of IFNβ might contribute to, but may not fully explain, the marked overproduction of type 1 IFN-inducible transcripts and proteins seen in DM muscle and blood.
Statistics from Altmetric.com
Dermatomyositis (DM) is an autoimmune disease with prominent muscle and skin involvement. DM muscle,1 2 blood2 and skin3 contain markedly increased amounts of many type 1 interferon (IFN)-inducible transcripts and proteins.1 4 The mechanisms driving the overproduction of type 1 IFN-inducible transcripts and proteins in DM muscle remain unknown. Differential presence of type 1 IFNs themselves has not been demonstrated, either at the level of transcript or protein. Here, we measured IFNα, IFNβ and IFNω protein in blood samples from patients with myositis and healthy volunteers and correlated these measurements with the abundance of type 1 IFN-inducible gene transcripts in these same blood samples, and with functional activity in a bioassay. We found that IFNβ was uniquely associated with DM.
Patients and methods
Patients and blood samples
One hundred and twenty-nine blood samples were collected, from 93 patients with muscle diseases and from 36 healthy volunteers. Patients fulfilled criteria for definite or probable polymyositis (PM; N=26; mean age 60) or DM (N=26; mean age 48),5 inclusion body myositis (IBM; N=22; mean age 67),6 or had other muscle diseases (muscular dystrophy N=6, toxic myopathy N=4, idiopathic N=3, or myasthenia gravis N=6). Healthy volunteers (mean age 41) stated they were healthy, had no medical problems and had had no cold or flu-like illness or vaccinations within 3 months. The patients with myositis comprised 48 women and 26 men and the healthy volunteers, 24 women and 12 men. Patients with DM and PM were stratified as untreated (having never received immunomodulatory treatment; DM N=7, PM N=12), minimally treated (prednisone dose ≤15 mg/day or treatment duration ≤7 days and no other immunomodulatory agents; DM N=4, PM N=2), or chronically treated (any other prednisone dose or duration, or other immunomodulatory agent; DM N=15, PM N=12). Patients with IBM were untreated. All patients had predominantly muscle disease.
Patients provided informed consent for research studies, approved by our institutional review boards. Blood was collected in EDTA tubes and plasma and buffy coats were separated in clotted tubes for serum and PAXgene tubes (PreAnalytix GmbH, Hombrechtikon, Switzerland) for some of the RNA experiments. All samples were stored at −80° C in multiple aliquots.
All samples (N=129) were assayed for IFNα, IFNβ and IFNω protein by ELISA; 123 samples were assayed for their ability to stimulate type 1 IFN-inducible gene transcription in a functional bioassay. Microarray gene expression profiling was performed on a subset (N=70) before performance of the other studies, some of these data being previously reported.2
IFN ELISAs
Concentrations of type 1 IFNs in serum or plasma were measured using the following ELISAs according to the manufacturer's protocol: IFNα catalogue No 41110 (high-sensitivity protocol), IFNβ catalogue No 41415 and IFNω catalogue No 41395, all from PBL InterferonSource, Piscataway, New Jersey, USA. Assays were extended to concentrations below the lower limit of detection through additional standards. Spike-in recovery experiments using IFNβ showed that in EDTA-preserved plasma, a mean of 89% of low spike (5 pg/ml) and 96% of medium spike (25 pg/ml) IFNβ was detected by the ELISA. All samples were tested in duplicate on the same plate, in adjacent wells. Standard curves were repeated in each batch of 96 wells assayed.
Induction of type 1 IFN-inducible gene transcription in a bioassay
A 293T-ISRE (IFN-stimulated response element) cell line was cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, glutamine and sodium pyruvate. Approximately 10 000 cells were seeded into each well of a 96-well plate and incubated overnight at 37°C. Cells were then treated with serum and plasma of various concentrations for 9 h. Supernatants were removed and cells lysed in Bright-Glo Luciferase Assay System (Promega Corporation, Madison, Wisconsin, USA) for 3 min at room temperature. Luminescence was measured using a luminometer (SpectraMax L; Molecular Devices, Sunnyvale, California, USA). Activity of type I IFN was calibrated as the equivalent ISRE induction by recombinant human IFNα 2a (PBL Interferon Source) in standard curves performed from each plate. Assay sensitivity was 15 pg/ml of IFNα.
Both serum and plasma were found to have inhibitory effects in this bioassay. The effects were present at all concentrations (5%, 25%, 40% and 50%) tested and greatest in plasma at highest concentrations. We therefore performed all assays of plasma using standard curves constructed from IFNα standards added to healthy volunteer plasma and separately for serum using standards added to healthy volunteer serum.
Blood microarray gene expression profiling
Microarray gene expression profiling was performed on RNA from the same blood specimen for 70 of the 129 samples assayed by ELISA (DM N=24, PM N=12, IBM N=15, other myopathy N=7, normal N=12) using Affymetrix (Santa Clara, California, USA) U133 Plus 2.0 microarray assays measuring approximately 55 000 probesets representing the entire human genome as previously described.2 A gene composite score representing the median fold change of the four most highly expressed transcripts in DM and PM blood2 was calculated for each blood sample.
Results
High sensitivity and reproducibility of IFN protein subtype ELISAs
We verified the high sensitivity of the ELISAs used in these studies (figure 1) and extended these assays to detection limits of 3.13 pg/ml for IFNα, 1.15 pg/ml for IFNβ and 2.40 pg/ml for IFNω. These ELISAs demonstrated high reproducibility in paired measurements of all 129 samples (r2=0.99 for IFNα, 0.98 for IFNβ and 0.93 for IFNω).

ELISA performance. (Left) Standard curves. Performance remains linear at least to 3.13 pg/ml for IFNα, 1.15 pg/ml for IFNβ and 2.4 pg/ml for IFNω. (Right) High reproducibility for duplicate measurements.
Healthy volunteers: frequent artefactual detection of IFNα and IFNω protein
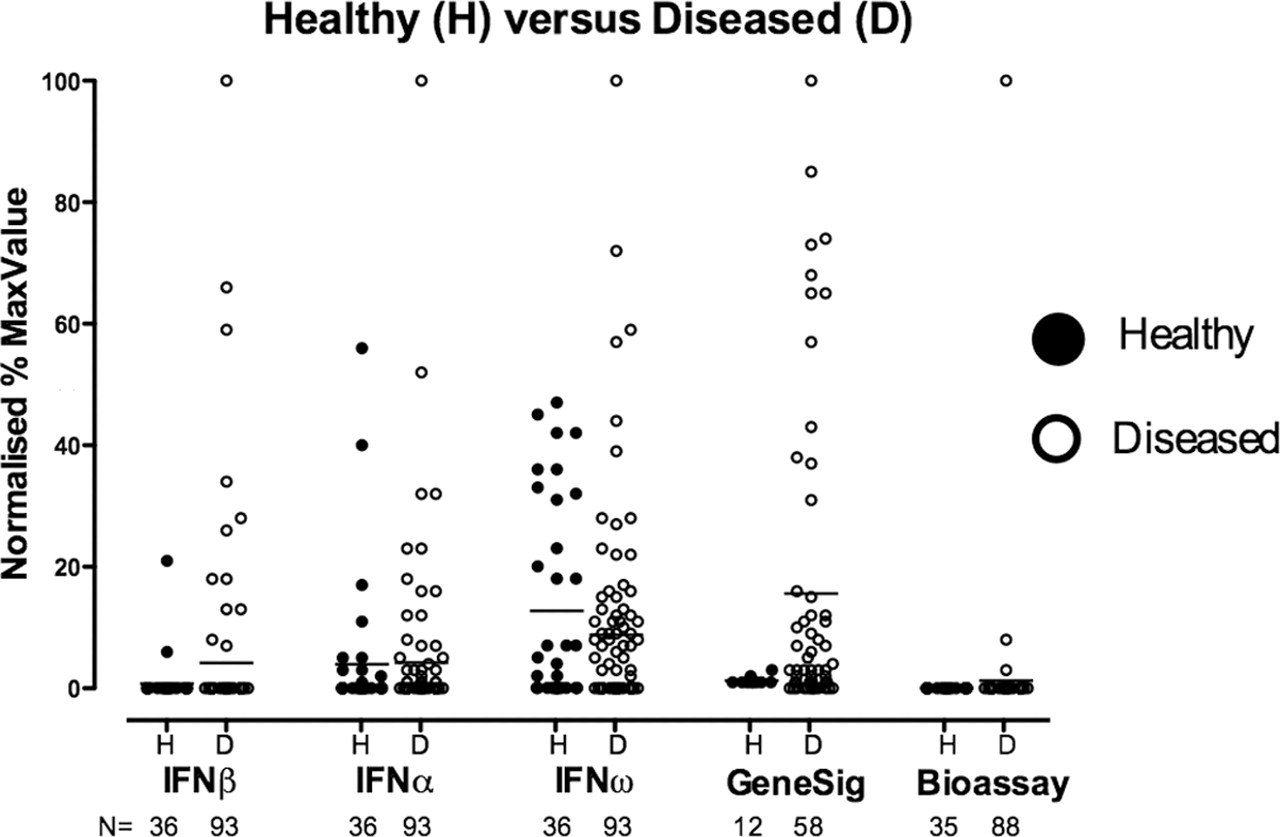
IFNα was detected in 31% and IFNω in 53% of healthy volunteer (N=36) serum or plasma samples. Some healthy volunteer samples contained high IFNα concentrations, up to 835 pg/ml, with 28% of volunteer samples having >15 pg/ml of IFNα and 36% having >15 pg/ml of IFNω (figure 2). None of 35 of these samples tested demonstrated IFN-inducible responses in a bioassay of IFN activity, including 10 samples with IFNα concentrations greater than the bioassay's 15 pg/ml sensitivity. Similarly, microarray gene expression profiling was performed on the same blood samples from 12 of the healthy volunteers. None of these had increased peripheral blood mononuclear cell (PBMC) type 1 IFN-inducible gene transcription, including three samples with high ELISA-measured IFNα concentrations of 167 pg/ml, 252 pg/ml and 835 pg/ml.

Comparison of five assays in healthy (N=36) and diseased (N=93) blood samples. Three ELISA protein assays (interferon (IFN)β, IFNα, IFNω), a microarray gene expression signature (GeneSig) and a functional reporter bioassay were used for healthy volunteers, with number of samples shown for each assay. Comparison of healthy and diseased samples in each assay shows considerable overlap of healthy volunteer and diseased sample IFNα and IFNω protein measurements but distinct differences in IFNβ protein and gene expression signatures. Bioassay positivity, although only present in diseased samples, was uncommon.
To determine if the ELISA-measured IFNα values were false positive artefacts due to assay-interfering biological substances such as anti-heterophile antibodies in these samples, we performed dilution experiments with sample buffer. Rather than linear reduction in ELISA IFNα signal as would be expected if such a signal indeed reflected true IFNα concentration,7 we found instead that for the four healthy volunteer samples tested, all with >100 pg/ml, dilution 1:2 led to a complete loss of signal (<6.5 pg/ml) (figure 3A).
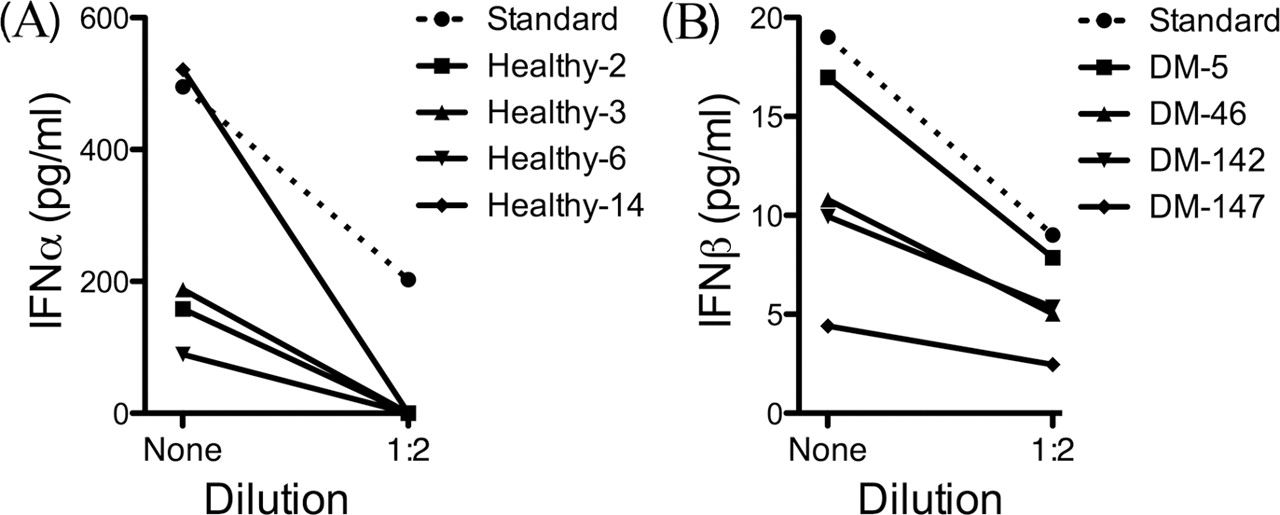
Artefactual interferon (IFN)α detection in healthy volunteers; accurate IFNβ detection in diseased samples. (A) High-levels of an ELISA-detected IFNα signal in four healthy volunteers (Nos 2, 3, 6 and 14; range of detected IFNα signal 89–521 pg/ml) become undetectable (<6.25 pg/ml) at 1:2 dilution, in contrast to recombinant IFNα spike in samples (Standard) which instead dilute linearly. (B) Detectable IFNβ signal in four dermatomyositis (DM) samples (Nos 5, 46, 142 and 147; range of detected IFNβ signal 4.4–17.6 pg/ml) reduces exactly by a half at 1:2 dilution. Mean of two replicates shown for all samples.
In contrast, in healthy volunteers IFNβ was infrequently detected (6% of healthy volunteers), with all 36 samples testing below the assay limit of 1.15 pg/ml except one each with 1.2 pg/ml and 4.1 pg/ml.
Of the five assays examined in this study, only IFNβ protein and type 1 IFN-inducible gene expression distinguished diseased samples from healthy volunteer samples (figure 2).
IFNβ but not IFNα or IFNω protein was highly associated with DM
IFNβ but not IFNα or IFNω protein concentration was associated with DM (p=0.0004; figure 4). No other association was present between a type 1 IFN and a muscle disease subtype. Detectable IFNβ (>1.15 pg/ml) was present in 35% (9/26) of patients with DM compared with 6% (3/48) of other inflammatory myopathies (IBM+PM), which did not differ from 6% (2/36) of healthy volunteers. Stratification by treatment status demonstrated a stronger association between IFNβ and DM (figure 4B). Detectable IFNβ was present in 64% (7/11) of samples from untreated or minimally treated (prednisone dose of ≤15 mg/day or treatment duration ≤7 days) patients with DM and 13% (2/15) of chronically treated DM samples, but 7% of patients with untreated or minimally treated PM (1/14), 0% of chronically treated PM (0/12) and 9% of untreated IBM (2/22). The concentrations of IFNα, IFNβ and IFNω did not differ in IBM and PM compared with healthy subjects.

Association of blood interferon β (IFNβ) with dermatomyositis (DM). (A) IFNβ was significantly associated with DM but not with inclusion body myositis (IBM), polymyositis (PM), or non-inflammatory myopathies (OtherMyo) in 93 muscle-diseased and 36 healthy volunteer samples (p=0.0004 Kruskal–Wallis analysis of variance (ANOVA) of five groups). (B) Detectable IFNβ was present in 64% (7/11) of untreated or minimally treated DM patients (MinTx), who had higher IFNβ concentrations than patients with chronically treated (Tx) DM (p=0.01, Mann–Whitney). (C, D) No differences in IFNα or IFNω among myositis subtypes were present (p=0.20 and 0.33, Kruskal–Wallis ANOVA of five groups). Bar represents mean value in all panels.
To ensure that IFNβ detection was not artefactual, a dilution experiment, as had been performed for IFNα in four healthy volunteer samples, was similarly performed for four DM samples. When diluted 1:2, these samples all showed expected 50% lower IFNβ levels (figure 3B).
Correlation of IFNβ but not IFNα or IFNω protein with type 1 IFN-inducible gene expression in the same blood samples
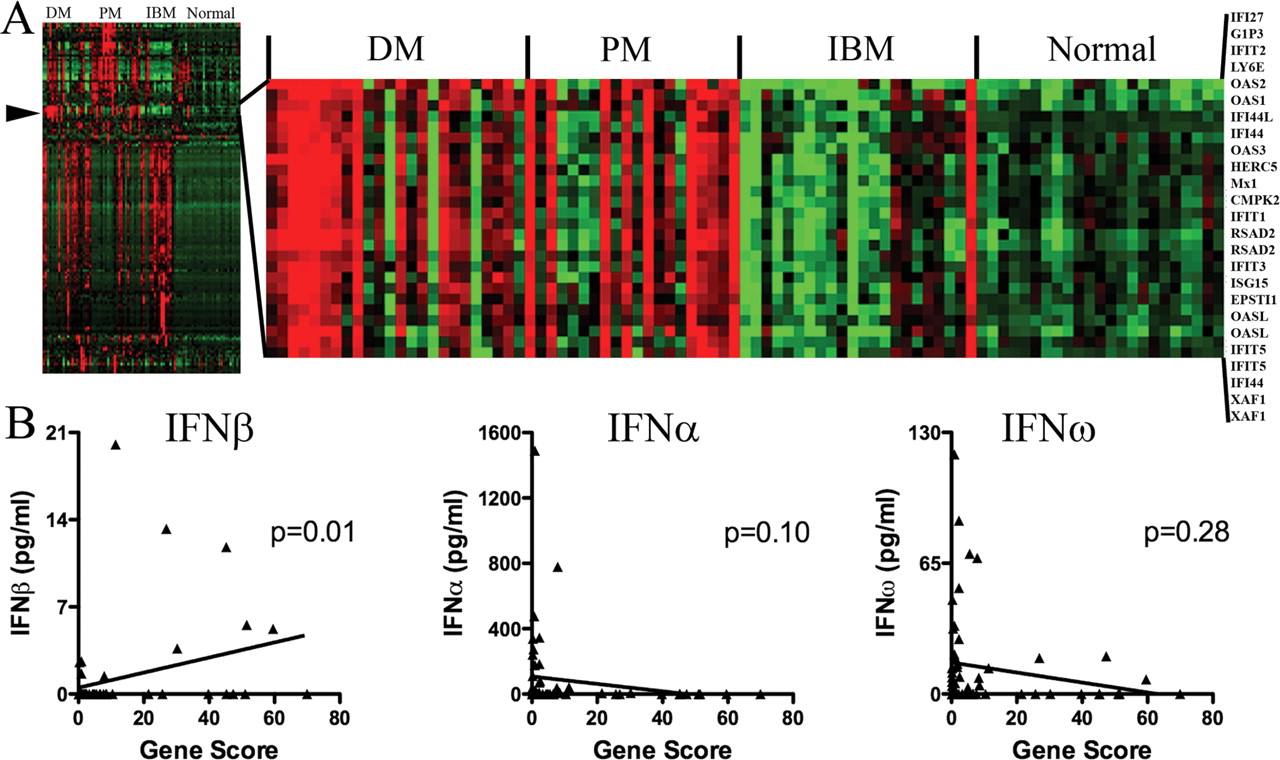
Type 1 IFNs have the ability to induce the transcription of many type 1 IFN-inducible genes. For 70 blood samples, we separated serum or plasma from PBMCs and compared the concentration of type 1 IFNs in the serum or plasma with the expression of type 1 IFN-inducible genes in the PBMCs. High expression of type 1 IFN-inducible transcripts (gene composite score of greater than fivefold) was present in 54% (13/24) of DM, 67% (8/12) of PM and 5% (1/22) of IBM samples (figure 5A).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlation of interferon (IFN)β but not IFNα or IFNω protein with type 1 IFN-inducible gene expression in the same blood samples. (A-Left) Unsupervised clustering of gene expression profiles of 78 blood samples disclosed a cluster (arrowhead) of highly expressed genes in dermatomyositis (DM). A total of 1656 genes (one per row) is shown, with red high expression, black neutral and green low expression (A-Right). This cluster is expanded and shows intense expression in DM and polymyositis (PM) of 25 probe sets all of which are type 1 IFN-inducible genes. (B) Type 1 IFN protein levels in serum or plasma compared with type 1 IFN-inducible transcripts in peripheral blood mononuclear cells from the same blood samples. IFNβ but not IFNα or IFNω proteins showed linear correlation with type 1 IFN-inducible transcript abundance (Gene Score).
IFNβ (r= 0.32; p=0.01), but not IFNα (r= −0.22; p=0.10) or IFNω (r= −0.14; p=0.28; Spearman rank correlation for all), protein concentrations correlated with high type 1 IFN-inducible gene expression in the same blood samples (figure 5B). Marked elevation of the gene composite score (>10-fold) in eight specimens, all with DM, was accompanied by elevated IFNβ in 75% of these; none of these specimens had elevated IFNα or IFNω protein concentrations. Increased IFN-inducible gene expression was not accompanied by increased concentrations of type 1 IFNs in 75% of samples.
Diseased and healthy volunteer samples with high levels of IFNα and IFNω do not induce type 1 IFN signalling in a functional bioassay
Of 123 samples tested in a functional bioassay for their ability to induce type 1 IFN signalling in a reporter assay, only three (2%) had detectable IFN-inducing ability above the assay sensitivity (the equivalent of approximately 15 pg/ml of IFNα) (figure 2). Two of these were DM samples and one a PM sample. All other samples, many with high levels of IFNα (up to 1491 pg/ml) and IFNω (up to 199 pg/ml), had no activity in the functional assay. The sample with the highest IFNβ protein concentration (20 pg/ml) had the highest bioactivity (1239 pg/ml of IFNα equivalence), while other samples with IFNβ (up to 13 pg/ml) had no detectable bioactivity, though such activity was below our assay's detection threshold of 15 pg/ml.
Discussion
DM is distinguished from other myopathies by the marked overabundance of type 1 IFN-inducible transcripts and proteins in muscle1 4 and the presence of endothelial tubuloreticular inclusions, a known biomarker of type 1 IFN exposure.8,–,10 The molecules and mechanisms driving these molecular changes in people with DM remain unknown (reviewed by Greenberg 11).
The current studies demonstrate an association between IFNβ in blood and DM. Whereas high levels of IFNα and IFNω were commonly detected in blood samples from healthy volunteers and patients with myositis, the detection of IFNβ even at comparatively low concentrations was highly specific for DM. Accordingly, these findings support the hypothesis that in some patients with DM IFNβ might drive production of type 1 IFN-inducible transcripts and proteins in muscle and blood. The concentrations of IFNβ detected in 64% of patients with untreated or minimally treated DM can induce a molecular picture in cultured human skeletal muscle similar to that present in human DM biopsy specimens.4 IFNβ concentration in muscle could be considerably higher than in blood, given accumulation of plasmacytoid dendritic cells, immune cells specialised in the production of type 1 IFNs, in DM muscle1 12 and the marked enrichment of type 1 IFN-inducible transcripts in muscle compared with blood.2
We found little activity in a functional bioassay of the diseased samples, in only three samples, though the assay used had less sensitivity for detection of type 1 IFNs than ELISA. In a study of juvenile DM, serum from approximately 4 of 18 (22%) untreated patients had higher than normal (compared with age-matched healthy subjects) ability to stimulate type 1 IFN-inducible gene expression in a similar bioassay.13 These differences might be due to differences in assay sensitivity, disease acuity, treatment status or underlying biology. We also found high levels of ELISA-measured IFNα in healthy volunteers that failed to induce IFN signalling in bioassays, as a previous study reported.14 We confirmed that these ELISA values are artefactual, due to interfering biological molecules in the samples, addressing longstanding concerns in the field as to the nature of the high IFNα ELISA signal in the healthy population.
These studies have two important limitations. They do not clarify the mechanism of high type 1 IFN-inducible gene expression in many patients with myositis whose samples lacked detectable type 1 IFN proteins. Possibly, these ELISAs did not detect a biologically active type 1 IFN (some were not measured, such as IFNκ), or the PBMCs containing high abundance of type 1 IFN-inducible transcripts had recirculated from tissue containing higher concentrations of type 1 IFNs, or the IFN-inducible response was being sustained without ongoing type 1 IFN receptor signalling by type 1 IFNs. Although an association of IFNβ with DM suggests the possibility that IFNβ may drive tissue injury in DM, whether IFNβ definitively contributes to muscle injury in some patients with DM cannot be determined without selective neutralisation of IFNβ in people, which currently is not possible.
Acknowledgments
The authors thank MedImmune, LLC, Gaithersburg, MD for performing some of the gene expression experiments.
References
Footnotes
-
Funding Funded through Brigham and Women's Hospital neurology departmental funds and philanthropic contributions.
-
Competing interests Dr Jubin's employer manufactures an assay for the detection of interferon β.
-
Ethics approval This study was conducted with the approval of the Brigham and Women's Hospital and Children's Hospital Boston.
-
Provenance and peer review Not commissioned; externally peer reviewed.