Article Text

Abstract
Objectives Patients with systemic lupus erythematosus (SLE) have an ongoing interferon-α (IFN-α) production by plasmacytoid dendritic cells (pDCs). We investigated whether T cells can promote IFN-α production by pDCs.
Methods Human pDCs were stimulated with immune complexes (ICs) containing U1 small nuclear ribonucleic proteins particles and SLE-IgG (RNA-IC) in the presence of T cells or T cell supernatants. T cells were activated by anti-CD3/CD28 antibodies or in a mixed leucocyte reaction. IFN-α and other cytokines were determined in culture supernatants or patient sera with immunoassays. The effect of interleukin (IL) 3 and granulocyte-macrophage-colony-stimulating factor (GM-CSF) on pDCs was examined by the use of antibodies, and the expression of CD80/CD86 was determined using flow cytometry.
Results Activated T cells and supernatants from activated T cells increased IFN-α production by >20-fold. The stimulatory effect of T cell supernatants was reduced after depletion of GM-CSF (81%) or by blocking the GM-CSF receptor (55%–81%). Supernatant from activated T cells, furthermore, increased the frequency of CD80 and CD86 expressing pDCs stimulated with RNA-IC from 6% to 35% (p<0.05) and from 10% to 26% (p<0.01), respectively. Activated SLE T cells enhanced IFN-α production to the same extent as T cells from healthy individuals and a subset of patients with SLE had increased serum levels of GM-CSF.
Conclusions Activated T cells enhance IFN-α production by RNA-IC stimulated pDCs via GM-CSF and induce pDC maturation. Given the increased serum levels of GM-CSF in a subset of patients with SLE, these findings suggest that activated T cells may upregulate type I IFN production in SLE.
- Systemic Lupus Erythematosus
- T Cells
- Cytokines
Statistics from Altmetric.com
Introduction
Systemic lupus erythematosus (SLE) is an autoimmune disease characterised by the production of autoantibodies against nuclear antigens, immune complex (IC) formation and an ongoing production of type I interferon (IFN), for example, IFN-α.1 ,2 IFN-α is mainly produced by plasmacytoid dendritic cells (pDCs), which can be activated by ICs containing autoantibodies in combination with RNA or DNA derived from apoptotic or necrotic cells.3 Such interferogenic ICs are internalised to the endosomes via FcγRIIA and the nucleic acids interact with Toll-like receptor (TLR)7 or TLR9, resulting in gene transcription and protein synthesis.4 The produced IFN-α affects the immune system in a number of ways, including polarisation of naïve T helper (Th) cells to Th1 cells,5 restriction of development of Th26 and Th177 cells and expansion and activation of cytotoxic CD8+T cells.8 ,9 Furthermore, IFN-α prevents activated T cells from undergoing apoptosis10 and promote the development of long-lived central memory CD4+ and CD8+ T cells.11 ,12 In addition, IFN-α enhances B-cell differentiation, increases antibody production and promotes Ig class switch and maturation of dendritic cells.13 ,14 Thus, once pDCs are activated, they strongly promote the adaptive immune response.15 Much less is known about the effects on pDCs by the different adaptive immune cells. We previously demonstrated that B cells16 and NK cells17 ,18 enhance IFN-α production by pDCs stimulated with RNA-IC, but the possible role of T cells in the response of pDCs to interferogenic IC is unknown. Given the important role of T cells in the pathogenesis of SLE19 ,20 we investigated the capacity of T cells to modulate IFN-α production by RNA-IC stimulated pDCs.
Patients, materials and methods
Patients and healthy individuals
Patients (n=58) were recruited from the Rheumatology Unit at Uppsala University Hospital, Sweden, and fulfilled four or more of the American College of Rheumatology (ACR) classification criteria for SLE.21 The disease activity was determined using Systemic Lupus Erythematosus Disease Activity Index 2000 (SLEDAI-2000), where complement levels and anti-double-stranded DNA antibodies were excluded.22 Cumulative disease damage was measured using the Systemic Lupus International Collaboration Clinics/American College of Rheumatology Damage Index (SLICC/ACR DI).23 Demographic and clinical characteristics of the patients included in the serum granulocyte-macrophage-colony-stimulating factor (GM-CSF) analysis (n=51) are given in table 1. The patients participating in the T cell activation experiment (n=7) were all women (average age 38 years); one had a mild flare with a modified SLEDAI score of 1 (leucopenia). Treatment included hydroxychloroquine (n=6), low dose of prednisone (n=3), mycophenolate mofetil (n=1) and azathioprine (n=1). Healthy individuals and blood donors were recruited at the Department of Medicine or the Department of Clinical Immunology and Transfusion Medicine, both at Uppsala University Hospital. The study was approved by the local ethic committee, and all patients and healthy controls gave their informed consent.
Demographic and clinical characteristics of the patients with SLE included in the serum-GM-CSF analysis
Cell isolation
Peripheral blood mononuclear cells (PBMCs) were prepared by Ficoll-Hypaque (GE Healthcare) density-gradient centrifugation of blood donor buffy coats or from EDTA blood. CD3+, CD3+CD4+ and CD3+CD8+ T cells, myeloid dendritic cells (mDCs) and pDCs were isolated by negative selection (Miltenyi Biotec, Bergisch Gladbach, Germany). PBMCs (2×105) or pDCs (25×103) and T cells (as indicated) were cultured in 0.1 mL volumes in duplicates in 96-well plates for 20 h at 37°C with 5% CO2 as described previously.18
IFN-α inducers
To generate IFN-inducing ICs, total IgG was purified from the serum of a patient with SLE, containing autoantibodies to SmB, SmD, RNP-A, RNP-C, ribosomal P antigen, histone and dsDNA, by protein G chromatography.24 U1 snRNP particles purified from HeLa cells were a kind gift from Dr Gert Weber at Free University of Berlin, Germany.25 The U1 snRNP particles and SLE-IgG were used together in cell cultures at final concentrations of 2.5 µg/mL and 1 mg/mL, respectively. The phosphorothioate oligonucleotide ODN2216 (CyberGene, Stockholm, Sweden) and ultraviolet light-inactivated HSV type1 were used at concentrations of 3 µg/mL and 10% (v/v), respectively.24
T cell activation and mixed leucocyte reaction
T cells were activated using Dynabeads CD3/CD28 (Life Technologies) or in a mixed leucocyte reaction (MLR); see online supplementary information.
Blocking of GM-CSF and IL-3 in T cell supernatants, immunoassays, cell stimulation and flow cytometry
See online supplementary information.
Statistical methods
Differences between groups of paired measurements were analysed by Wilcoxon matched-pairs signed-rank test (GraphPad Prism V.6.0, San Diego, California, USA). For non-paired observations, categorical variables were compared with χ2 test and continuous variables by Mann–Whitney U Test (Statistica 12, Statsoft, Tulsa, Oklahoma, USA). p Values <0.05 were considered significant.
Results
Activated T cells enhance IFN-α production by pDCs stimulated with RNA-IC
To investigate whether T cells affect IFN-α production by RNA-IC stimulated pDCs, we cultured T cells together with pDCs in the presence of anti-CD3/CD28 or isotype-coated beads. Addition of CD3/CD28-activated T cells caused a >20-fold increase in IFN-α production compared with addition of non-activated T cells, which had a small effect (figure 1A). A similar increase in IFN-α production was seen when activated T cells were co-cultured with pDCs stimulated with ODN2216 or HSV (figure 1B, C). Both activated CD4+ and CD8+ T cells increased IFN-α production by RNA-IC stimulated pDCs (figure 1D, E). The increased level of IFN-α was also seen in cultures of PBMCs stimulated with RNA-IC and anti-CD3/CD28 (figure 1F). Intracellular staining and flow cytometric analysis showed that pDCs, but not T cells, produced IFN-α in the cell cultures (results not shown).
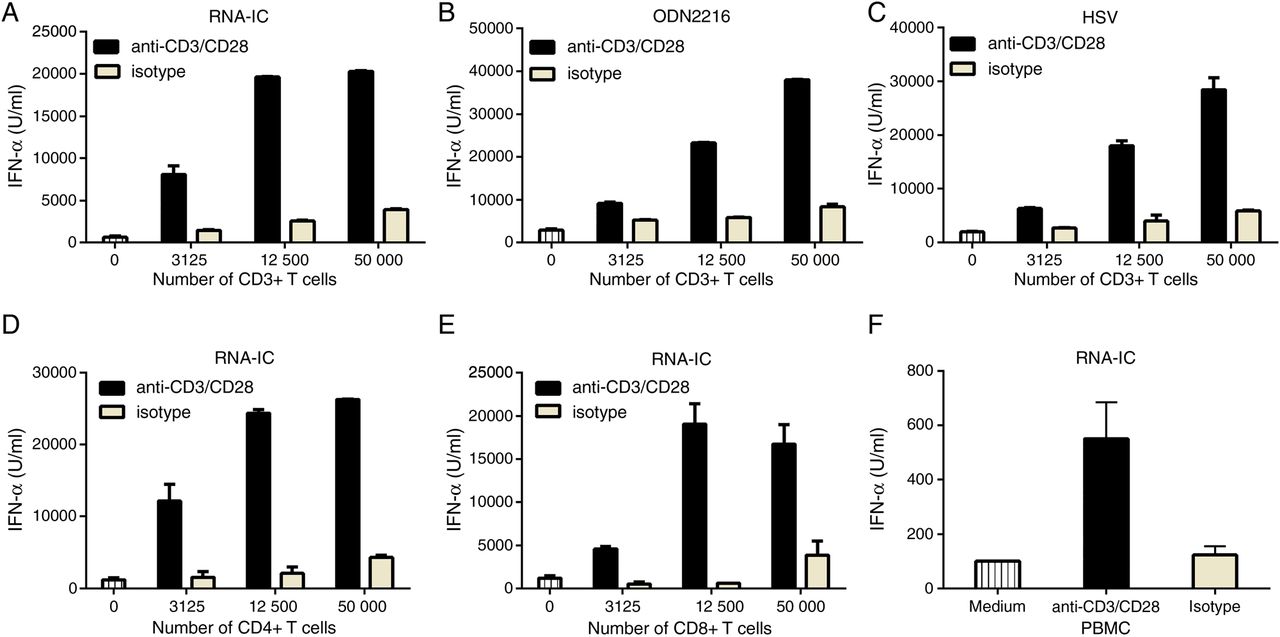
Activated T cells enhance interferon-α (IFN-α) production by plasmacytoid dendritic cells (pDCs). (A–E) pDCs (25×103/well) were cultured alone or together with T cells in the presence of anti-CD3/CD28 or isotype beads and the indicated stimuli. (F) Peripheral blood mononuclear cells (PBMCs) (2×105/well) and RNA-immune complex (IC) were cultured in the presence of anti-CD3/CD28 or isotype beads. Data are shown for (A–C) total CD3+ T cells, (D) CD4+ T cells, (E) CD8+ T cells and (F) PBMCs alone. IFN-α was measured after 20 h by an immunoassay. Bars show the mean±SD of duplicate wells. A, B, C and F are representative of three experiments. D and E were performed once.
Soluble factors enhance IFN-α production by pDCs stimulated with RNA-IC
In order to determine whether the enhancing effect of T cells on IFN-α production is mediated via soluble factors, we added supernatants from anti-CD3/CD28-treated T cells to RNA-IC activated pDC cultures. Such supernatants increased IFN-α production in a dose-dependent manner (figure 2A). Supernatants from anti-CD3/CD28-treated CD4+ as well as CD8+ T cells increased IFN-α production by >15-fold, compared with pDCs cultured with supernatants from T cells treated with isotype IgG (figure 2B). Time-course studies showed that IFN-α could be detected in the pDC cultures 4–6 h after stimulation, with a maximum at 24 h (figure 2C).

Supernatant from activated T cells enhance interferon-α (IFN-α) production by plasmacytoid dendritic cells (pDCs) stimulated with RNA-containing immune complexes (RNA-ICs). (A) Titration of volume supernatant from CD3+ T cells. (B) Effect of supernatant from CD3+, CD3+CD4+ or CD3+CD8+T cells cultivated with anti-CD3/CD28 or isotype IgG. (C) Time course of cumulative IFN-α production by pDCs in the presence of supernatant from anti-CD3/CD28 beads activated CD3+CD4+ or CD3+CD8+ T cells. (D) Mixed leucocyte reaction (MLR); CD3+ T cells and myeloid dendritic cells (mDCs) from different individuals (MLR) or from the same individual (auto-MLR) were cultured together or separately for 6 days. The supernatant was added to cultures (50% (v/v)) with pDCs and RNA-IC. Interferon-α (IFN-α) immunoassay was performed after 20 h (except in C). A–D are the mean±SD of duplicate wells. A, B and D are representative of three experiments, and C was performed once.
We next examined if supernatants from T cells activated in an MLR also could promote the IFN-α response. When MLR supernatants were added to the RNA-IC-stimulated pDCs, a 15-fold increase in IFN-α production was noted compared with cultures with pDCs stimulated with RNA-IC alone (figure 2D). Supernatants from an autologous MLR, or from CD3+ T cells or mDCs cultured separately did not affect IFN-α production (figure 2D).
In summary, activated CD4+ and CD8+ T cells produce soluble factor(s) that strongly enhance IFN-α production by pDCs stimulated with RNA-IC.
GM-CSF and IL-3 enhance IFN-α production by pDCs stimulated with RNA-IC
To identify the soluble factor(s) responsible for the increased IFN-α production we initially analysed the cytokine profile in activated T cell supernatants. High concentrations of interleukin (IL)-2 and GM-CSF, but low levels of IL-3 and IL-5, were detected (figure 3A). Next, we added the cytokines at the concentrations found in the T cell supernatants to the RNA-IC stimulated pDC cultures. Both GM-CSF and IL-3 had a strong stimulatory effect on the IFN-α response, and a combination of both cytokines increased IFN-α production as much as the activated T cell supernatants (figure 3B). A dose–response relationship was seen for both GM-CSF and IL-3 and notably, at similar concentrations, IL-3 was as potent as GM-CSF in enhancing IFN-α production (figure 3C). Depletion of GM-CSF from the supernatants significantly reduced the stimulatory effect (p<0.05) by on average 81%. Combined depletion of both GM-CSF and IL-3 did not further reduce the stimulatory effect (figure 3D). Addition of monoclonal antibodies (mAbs) blocking GM-CSF, the shared β-subunit of the GM-CSF and IL-3 receptor (CD131) or the α-subunit of the GM-CSF receptor (CD116) to the RNA-IC stimulated pDC cultures, significantly reduced the stimulatory effect of activated T cell supernatants (92%, 81% and 55%, all p<0.05) (figure 3E). However, antibodies to IL-3 or the α-subunit of the IL-3 receptor (CD123) did not significantly reduce the stimulatory effect of the activated T cell supernatant (data not shown).

Granulocyte-macrophage-colony-stimulating factor (GM-CSF) and interleukin (IL)-3 enhance interferon-α (IFN-a) production by plasmacytoid dendritic cells (pDCs) stimulated with RNA-containing immune complexes (RNA-ICs). (A) Cytokine profile in the supernatants from the activated CD3+ T cells (average from 3 donors); IFN-γ, tumour necrosis factor α, GM-CSF, interleukin-2 (IL-2), macrophage inflammatory protein 1α (MIP-1α), MIP-1β, IL-3 and IL-5 measured by immunoassays. (B) Effect of cytokines added at concentrations given in (A), supernatant from anti-CD3/CD28 activated CD3+ T cells (T-sup) (25%(v/v)). (C) Effect of different concentrations of GM-CSF or IL-3. (D) Effect of supernatant from anti-CD3/CD28 activated CD3+ T cells (1% (v/v)) depleted of GM-CSF and/or IL-3. (E) Effect of antibodies blocking CD131,26 GM-CSF or CD116.44 (D-E) pDC stimulated with RNA-IC is set as 0% and pDC stimulated with RNA-IC in the presence of supernatant from anti-CD3/CD28 activated CD3+ T cells (1% (v/v)) is set as 100%. Data are mean±SD of duplicate wells. B and C are representative of three experiments.
Next, the concentration of different cytokines was analysed in the supernatant from RNA-IC stimulated pDCs cultured together with GM-CSF. Increased concentration of IL-6, IL-8, tumour necrosis factor α and MIP-1β was seen (see online supplementary figure S1). We then asked if the increased IFN-α production by GM-CSF was caused by an increase in the number of IFN-α-producing pDCs. Intracellular staining for IFN-α showed that addition of GM-CSF or activated T cell supernatant to the RNA-IC-stimulated pDCs increases the frequency, but not the median fluorescence intensity of IFN-α expressing pDCs (see online supplementary figure S2). In addition, no proliferation of RNA-IC stimulated pDCs could be detected following culture with GM-CSF for 20 h (data not shown).
Taken together, our results show that both IL-3 and GM-CSF can enhance IFN-α production of RNA-IC-stimulated pDCs, but GM-CSF seems to be more important in supernatants from activated T cells.
Cytokines from activated T cells enhance the expression of costimulatory molecules on pDCs
After activation and type I IFN production, the pDC has a number of different functions, which is reflected in the expression of new cell surface molecules. Therefore, we investigated the effect of activated T cells on the expression of costimulatory molecules on pDCs. Supernatants from activated T cells added to pDCs stimulated with RNA-IC increased the frequency of pDCs expressing CD80 and CD86 from 6% to 35% (p<0.05) and 10% to 26% (p<0.01), respectively (figure 4A, B). A similar increased frequency of CD80 and CD86 expressing pDCs was seen in the presence of GM-CSF and IL-3 (figure 4A, B). Addition of RNA-IC to pDCs cultured with GM-CSF and IL-3 resulted in an increased frequency of CD80 and CD86 (p<0.05) expressing pDCs (figure 4A, B). The increased frequency of pDCs expressing CD80 and CD86 was significantly (p<0.05) reduced when GM-CSF and IL-3 were depleted from the T cell supernatant (figure 4C, D). Thus, activated T cells increase the expression of the costimulatory molecules CD80 and CD86 on pDCs stimulated with RNA-IC via GM-CSF and IL-3.
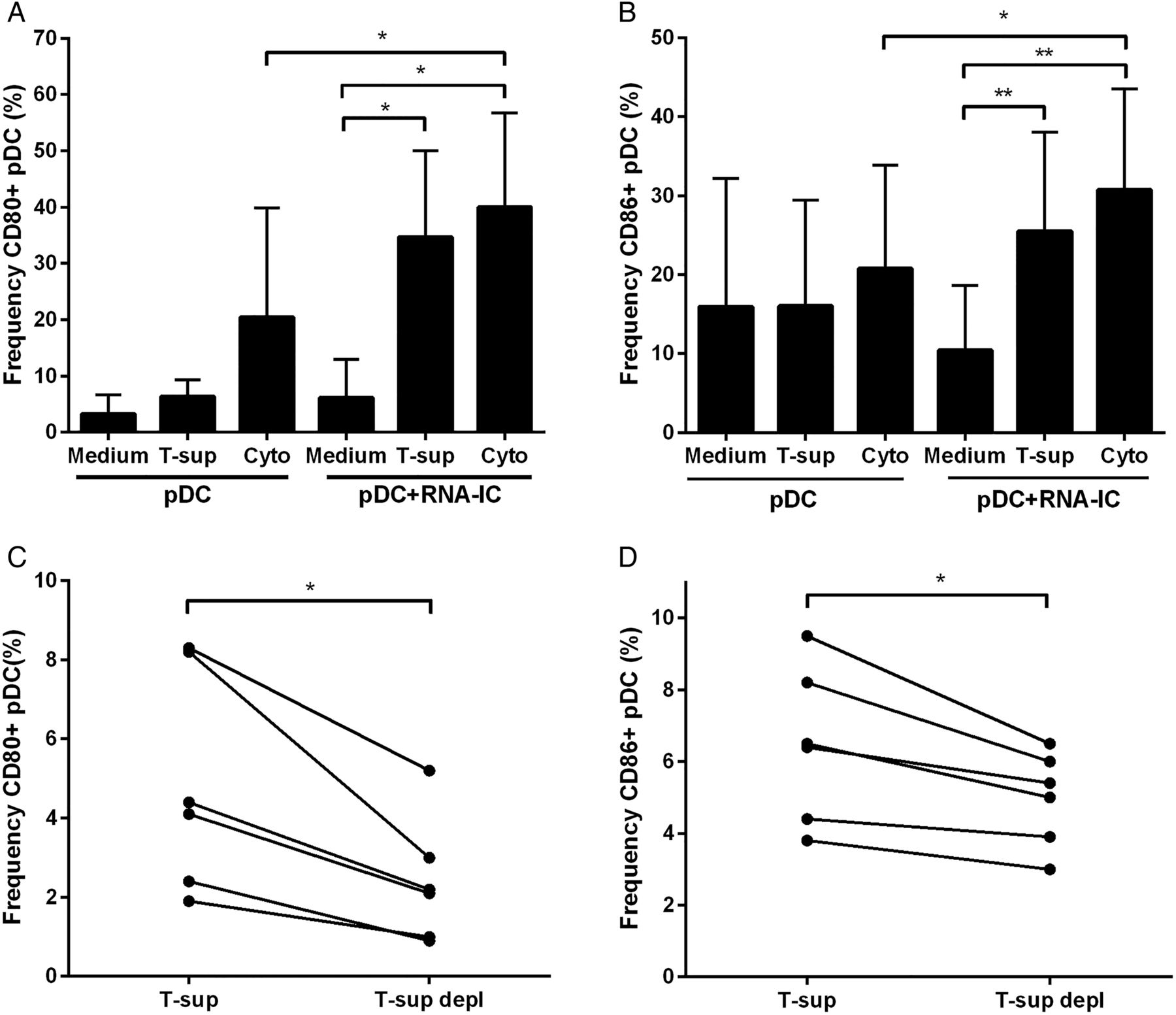
Supernatant from activated T cells and RNA-containing immune complexes (RNA-ICs) enhance the expression of CD80 and CD86 on plasmacytoid dendritic cells (pDCs). (A-B) Flow cytometric analysis of (A) CD80 and (B) CD86 expressing pDCs cultured 18 h with or without RNA-IC and supernatant from anti-CD3/CD28 activated CD3+ T cells (13% (v/v)) (T-sup) or granulocyte-macrophage-colony-stimulating factor (GM-CSF) (1 ng/mL) and interleukin (IL)-3 (0.25 ng/mL) (Cyto). (C-D) Flow cytometric analysis of (C) CD80 and (D) CD86 expressing pDCs, cultured 18 h with RNA-IC and anti-CD3/CD28 activated CD3+ T cell supernatant (T-sup) or activated CD3+ T cell supernatant depleted of GM-CSF and IL-3 (T-sup depl) (1% (v/v)). Bars show the mean±SD of A (n=7) and B (n=9) experiments.
T cells and serum from patients with SLE enhance IFN-α production
To determine whether T cells from patients with SLE also have the capacity to enhance IFN-α production, T cells from patients with SLE and age matched healthy controls were activated and cocultured with RNA-IC stimulated pDCs from a healthy blood donor. Activated SLE T cells could enhance IFN-α production to the same extent as T cells from healthy controls (figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Activated T cells from patients with systemic lupus erythematosus (SLE) and healthy individuals enhance the interferon-α (IFN-α)-producing capacity of plasmacytoid dendritic cells (pDCs). CD3+ T cells (25×103) were activated by anti-CD3/CD28 (aCD3/CD28) or isotype control IgG (Iso), and cocultivated with healthy donor pDCs in the presence of RNA-containing immune complexes (RNA-ICs). IFN-α levels were measured after 20 h by immunoassay and the bars show the mean values.
Given the prominent effect of GM-CSF on the IFN-α response, we measured serum levels of GM-CSF in 51 patients with SLE. We found detectable levels of GM-CSF (>15 pg/mL) in 31% of the patients, and this group was characterised by increased SLEDAI score (p<0.05) and a higher SLICC/ACR-DI (p<0.05) compared with patients with undetectable levels of s-GM-CSF (table 1). No association could be seen between the level of s-GM-CSF and any of the ACR criteria or the level of s-IFN-α. Next, sera from nine patients who had SLE with detectable levels of GM-CSF were added to cultures of pDCs stimulated with RNA-IC. Five of these sera strongly enhanced IFN-α production (>40%). Preincubation of these sera with a blocking anti-GM-CSF mAb resulted in significantly reduced IFN-α production compared with preincubation with isotype IgG (median reduction (range): 24% (16%–71%), p=0.02). Thus, GM-CSF in circulation is found in a subset of patients with SLE and the IFN-α promoting effect conferred by these SLE sera was reduced by neutralising GM-CSF.
Discussion
In the present study, we investigated the capacity of T cells to promote IFN-α production by pDCs in response to a stimulus considered to be one of the IFN-α inducers responsible for activation of the type I IFN system in SLE. Initially, we noted that unstimulated T cells together with pDCs and RNA-IC only marginally affected the IFN-α response by pDCs, indicating that RNA-IC per se could not trigger a T cell activation resulting in an increased pDC response. In contrast, when T cells were activated by either anti-CD3/CD28 antibodies or in an MLR, the T cells strongly enhanced IFN-α production by pDCs stimulated with RNA-IC. The stimulatory capacity after activation was similar for both CD8+ cytotoxic and CD4+ helper T cells. Hence, different T cell subsets have the capacity to markedly augment the pDC response to an SLE related TLR7 ligand. We also noted that activated T cells can enhance the IFN-α response to both a synthetic TLR9 ligand and HSV, the latter observation suggesting an important physiological role for T cells in promoting the pDC function. Taken together, our results show that T cells, in contrast to B cells16 and NK cells,18 require additional activation besides RNA-IC exposure before demonstrating an enhancing effect on IFN-α production by pDCs.
When the mechanisms behind the stimulatory effect of activated T cells on the IFN-α response were investigated, we noted that both GM-CSF and IL-3 demonstrated a strong stimulatory effect on pDCs. Notably, both these cytokines were equally potent at optimal concentrations, which might not be surprising as they share the same receptor β-subunit.26 However, GM-CSF was found at a much higher concentration than IL-3 in the supernatants and might therefore be of greater importance when T cells are activated. This assumption is supported by the observation that depletion of GM-CSF resulted in a significant reduction of the IFN-α response compared with depletion of IL-3 that did not inhibit IFN-α production. The importance of GM-CSF was further supported by the inhibitory effect of neutralising antibodies both to GM-CSF itself and to the α-chain and β-chain of the GM-CSF receptor. The intracellular signalling pathways by which GM-CSF in the setting of endosomal TLR signalling augment pDC IFN-α production is unknown. However, the PI3K- and MAPK-signalling pathways could be involved as they are activated by GM-CSF27 and have been implicated in IFN-α production in pDCs.28–30 Further, GM-CSF enhances IRF5 mRNA levels in monocytes,31 an important downstream transcription factor in the TLR signalling pathway.
Consequently, both GM-CSF and IL-3, which previously have been shown to promote the IFN-α response of PBMCs exposed to HSV,32 are of great importance also in promoting IFN-α production by pDCs stimulated with interferogenic ICs, but activated T cells mainly promote the IFN-α response via GM-CSF.
We noted that RNA-IC stimulated pDCs exposed to supernatants from activated T cells differentiate into a more mature state with increased expression of costimulatory molecules, which is in line with previous observations.33 Interestingly, RNA-IC by itself led only to a weak pDC differentiation, whereas the combination of RNA-IC and T cell supernatant provided a strong maturation signal to the pDCs. It is tempting to speculate that such a combined activation of the pDCs may occur in SLE, where pDCs will be exposed to both activated T cells and RNA-IC. This will eventually result in enhanced antigen-presenting capacity by the pDCs,34 which will promote the autoimmune process.
T cells from patients with SLE enhanced IFN-α production by pDCs in a similar way as T cells from healthy individuals. Given the presence of activated T cells in SLE35–40 one could perhaps expect a stronger enhancement of the IFN-α response by SLE-T cells compared with T cells from healthy individuals. One possible reason for the observed lack of difference could be that patients in the present study were all in clinical remission when the T cells were collected. Another explanation could be that all patients were taking immunosuppressive medication. However, despite immunosuppressive treatment, the SLE T cells, after activation, demonstrated a strong stimulatory effect.
Finally, we found that a subgroup of patients with SLE characterised by increased disease activity and disease damage had detectable levels of GM-CSF in serum. This is in line with previous findings of increased frequency of GM-CSF-secreting PBMCs in patients with SLE with active disease.41 Some of these GM-CSF-containing SLE sera enhanced IFN-α production when added to the RNA-IC-stimulated pDC cultures. We could also significantly reduce IFN-α production by the addition of antibodies blocking GM-CSF, suggesting a possible role for GM-CSF in the activation of the IFN system in SLE. Consequently, we suggest that blocking of GM-CSF could be one potential approach to reduce the elevated level of IFN-α in a subgroup of patients with SLE. However, the inhibitory effect was not complete, indicating other cytokines and chemokines also to be important in the pDC function and IFN-α production in SLE, which is perhaps not surprising given the large number of proinflammatory molecules present in patients with SLE.
In conclusion, our study shows that activated T cells via GM-CSF promote the RNA-IC-induced IFN-α production by pDCs. Given the elevated levels of GM-CSF41–43 in SLE, our results suggest that activated T cells could be involved in the process leading to the continuous type I IFN production seen in SLE.
Acknowledgments
The authors thank the patients and the healthy volunteers for their participation. The authors are grateful to Professor Angel Lopez (Hanson Center for Cancer Research, Adelaide, Australia) for the gift of the BION1 and 7G3 antibodies and Dr. Gert Weber for providing the U1snRNP complexes. They thank Charlotta Jakobsson, Anne Trönnberg and Lisbeth Fuxler for excellent technical assistance and Rezvan Kiani for collecting the blood samples.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Handling editor Tore K Kvien
Contributors DL designed the project, performed the experiments, analysed the data and wrote the article. M-LE and NH designed the project, performed the experiments, analysed the data and wrote the article. OB and KT performed the experiments and analysed the data. GA designed the project, analysed the data and wrote the article. LR designed the project, recruited the patients, analysed the data and wrote the article. All authors received and approved the final version of the manuscript.
Funding This project was funded by the Swedish Research Council for Medicine and Health (D0283001) and (A0258801), Swedish Rheumatism Foundation, King Gustaf V’s 80-year Foundation, Selander Foundation, Agnes and Mac Rudberg Foundation, Gustaf Prim Foundation, Torsten Söderberg Foundation, ALF-funding from Uppsala County Council and Uppsala University Hospital, the Knut and Alice Wallenberg Foundation (2011.0073) and COMBINE.
Competing interests None declared.
Ethics approval The Regional Ethical Review Board in Uppsala approved the study, and all participants gave their informed consent.
Provenance and peer review Not commissioned; externally peer reviewed.