


Abstract
(5R)-5-Hydroxytriptolide (LLDT-8) displays strong immunosuppressive activities both in vitro and in vivo in our previous studies. This study aims to investigate whether LLDT-8 has antiarthritic potential in a murine model of type II bovine collagen (CII)-induced arthritis (CIA) and to show the mechanism(s) of LLDT-8 action. DBA/1 mice were immunized with CII to induce arthritis and administered with LLDT-8. The severity of arthritis was evaluated according to the clinical score and joint damage. The effects of LLDT-8 on immune responses were determined by measurement of serum antibody levels, lymphocyte proliferation assay, cytokine assay, nitric oxide (NO) production, arginase activity assays, fluorescence-activated cell sorting analysis of splenic Mac-1+ cells, as well as polymerase chain reaction analysis for interferon-γ (IFN-γ)-related gene expression. We showed that LLDT-8 treatment significantly reduced the incidence and severity of CIA. The preventive and therapeutic effects of LLDT-8 are associated with 1) reduction of serum anti-CII immunoglobulin (Ig) G, IgG2a, and IgG1 levels; 2) inhibition of CII-specific lymphocyte proliferation, IFN-γ and interleukin-2 production; 3) blockade of gene expressions in IFN-γ signaling, including IFN-γ production pathways [signal transducer and activator of transcription (STAT) 1, T-box transcription factor, interleukin 12Rβ2, and STAT4] and IFN-γ-induced chemokine transcription [macrophage inflammatory protein (Mip)-1α, Mip-1β, regulated on activation normally T cell expressed and secreted, and inducible protein 10]; and 4) retardation of the abnormal increase of NO via IFN-γ/STAT1/interferon regulatory factor 1/inducible nitric-oxide synthase pathway and arginase activity. Moreover, the mRNA transcription of chemokine receptors was also suppressed [including C-C chemokine receptor (CCR) 1, CCR5, and C-X-C chemokine receptor 3]. In conclusion, our data suggest that the antiarthritic effect of LLDT-8 is closely related to the blockade of IFN-γ signaling. LLDT-8 may have a therapeutic value in the treatment of rheumatoid arthritis.
Tripterygium wilfordii Hook F. (TWHF) has been used as an herbal remedy to treat arthritis and other autoimmune disorders for centuries in China. Triptolide is the most potent immunosuppressive component in TWHF (Qiu and Kao, 2003). However, the therapeutic potential of triptolide is disrupted by its strong toxicity (Huynh et al., 2000). (5R)-5-hydroxytriptolide (LLDT-8) is a novel analog of triptolide that exhibits strong immunosuppressive activities but greatly reduced toxicities compared with its parent compound triptolide in our previous studies (Zhou et al., 2005, 2006). LLDT-8 could prevent graft-versus-host disease (Tang et al., 2005) and prolong the allogeneic cardiac graft survival (Tang et al., 2006).
Collagen-induced arthritis (CIA) is a well established murine model of human rheumatoid arthritis (RA) (Myers et al., 1997). The induction of severe chronic arthritis in naive mice requires type II collagen (CII)-specific activation of T cells and B cells and is associated with T helper (Th) 1-polarized immune responses. Interferon-γ (IFN-γ) and interleukin (IL)-2, derived from Th1 cells, predominate at the induction and acute phases of this disease (Choy and Panayi, 2001). There are two pathways participating in IFN-γ production signaling. One is the IFN-γ/signal transducer and activator of transcription (STAT) 1/T-box transcription factor (T-bet)/IFN-γ signaling; the other is the IL-12/STAT4/IFN-γ signaling (Grogan and Locksley, 2002).
Interferon regulatory factor (IRF)-1 is a transcription factor induced by IFN-γ and regulates interferon-related genes, including inducible nitric-oxide synthase (iNOS). The expression of iNOS leads to nitric oxide (NO) production by macrophages and other cells. Increased levels of NO in serum and synovial fluid have been reported in patients with RA (Farrell et al., 1992) and in animals with experimental arthritis (Stefanovic-Racic et al., 1994). Selective inhibition of the pathologically enhanced NO synthesis is a new experimental therapeutic approach in the treatment of inflammatory joint diseases (Stefanovic-Racic et al., 1994).
The ingress of leukocytes into sites of inflammation is crucial for the pathogenesis of arthritis and other inflammatory conditions. Chemokines and their receptors are involved in this process (Szekanecz and Koch, 2004). IFN-γ tightly regulates the expression of various chemokines, including macrophage inflammatory protein (Mip)-1α, Mip-1β, regulated on activation normally T-cell expressed and secreted (RANTES), and inducible protein 10 (IP-10) (Schroder et al., 2004). Their respective chemokine receptors, including C-C chemokine receptor (CCR) 5, C-X-C chemokine receptor (CXCR) 3, and CCR1, have also been implicated in inflammatory synovitis (Szekanecz and Koch, 2004). Blocking the expressions or functions of chemokine receptors will show therapeutic effects on arthritis.
TWHF suppressed the development of CIA in DBA/1 mice (Asano et al., 1998). Triptolide inhibited CIA in Lewis rat (Gu and Brandwein, 1998). However, the antiarthritic effects of extracts from TWHF are poorly understood. In this study, we showed that LLDT-8 significantly suppressed CIA through inhibiting IFN-γ production and its related signaling.
Materials and Methods
Mice. DBA/1 mice were housed in the animal facility of Shanghai Institute of Materia Medica. Male DBA/1 mice (7-8 weeks old, 20-22 g) were used in our experiments. The animals were housed under specific pathogen-free conditions. All the experiments were carried out according to the National Institutes of Health Guide for Care and Use of Laboratory Animals and were approved by the Bioethics Committee of Shanghai Institute of Materia Medica.
Induction and Assessment of Arthritis. CII was obtained from the Collagen Research Center (Tokyo, Japan) and dissolved in 0.1 M acetic acid. Male DBA/1 mice were immunized at the tail base with 125 μg of collagen emulsified in Freund's complete adjuvant (CFA) containing Mycobacterium tuberculosis strain H37Rv (Wako Pure Chemical Industries Ltd., Osaka, Japan) and boosted with the same preparations of collagen plus CFA 3 weeks later. The severity of arthritis was graded visually and expressed as the clinical score obtained by scoring each limb on a 0 to 4 severity grade and by summing the scores of four limbs [0 = normal; 1 = erythema or swelling of one or several digits; 2 = erythema and moderate swelling extending from the ankle to the mid-foot (tarsals); 3 = erythema and severe swelling extending from the ankle to the metatarsal joints; and 4 = complete erythema and swelling encompassing the ankle, foot, and digits, resulting in deformity and/or ankylosis; the maximum score for each animal is 16]. Day 0 was the time of booster immunization.

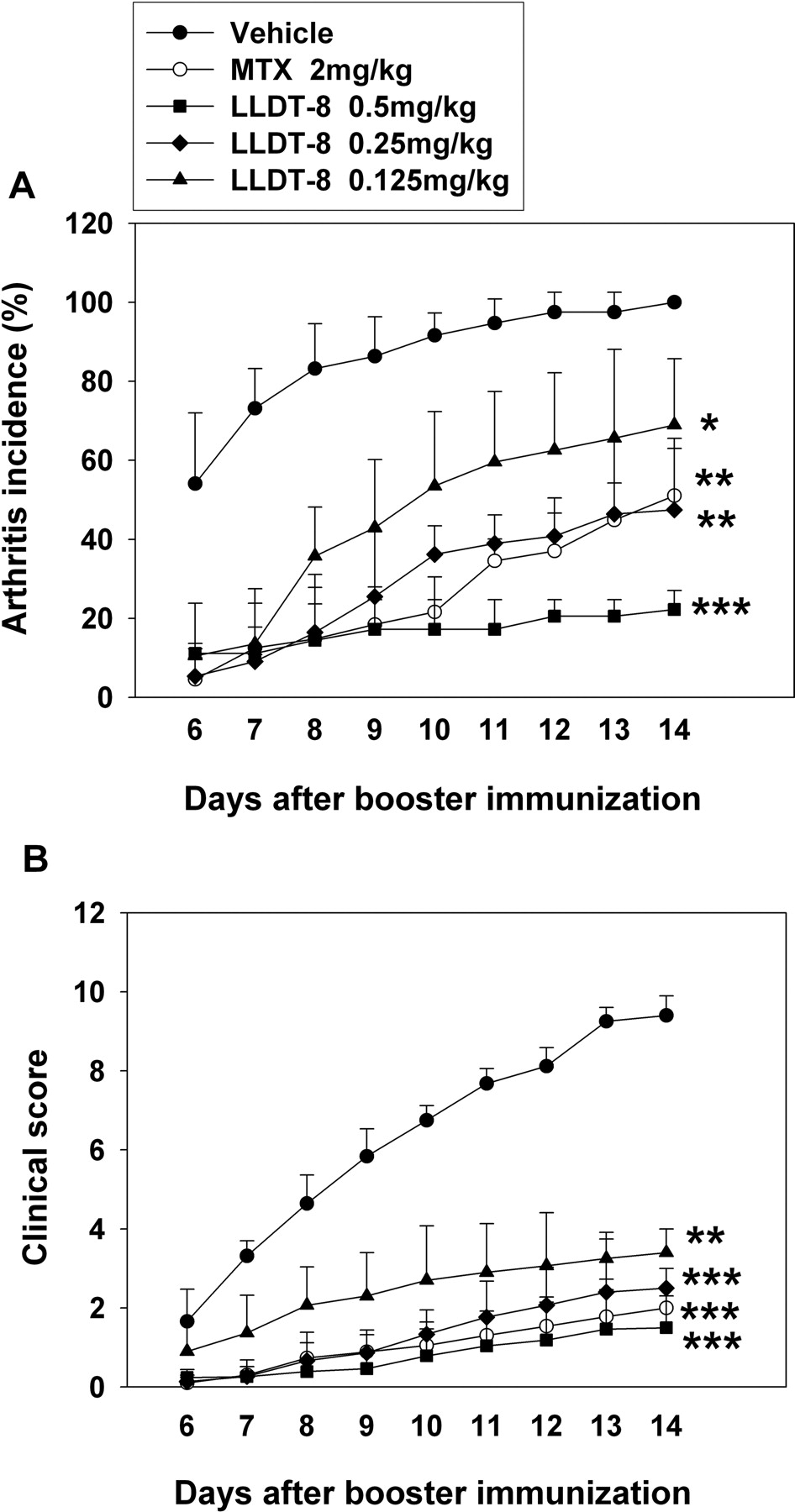
Administration of LLDT-8 reduced the incidence and severity of CIA in DBA/1 mice. DBA/1 mice were immunized with CII in CFA and boosted 3 weeks later to induce CIA. Mice were administered i.p. with vehicle, methotrexate at 2 mg/kg, or LLDT-8 at 0.125, 0.25, or 0.5 mg/kg once daily starting from 1 day before booster immunization for 2 weeks. The incidence (A) and severity (B) of CIA were evaluated. Data are mean ± S.E.M. of three separate experiments. *, p < 0.05, **, p < 0.01, ***, p < 0.001 versus vehicle-treated control mice; n = 12 mice/group.
Drugs and Administration. LLDT-8 (C20H24O7; MW = 376; 99% of purity) was synthesized from triptolide that was separated from TWHF (Zhou et al., 2005, 2006). The stock solution of LLDT-8 was prepared in dimethyl sulfoxide (Sigma, St. Louis, MO) at 300 mg/ml and further diluted with pathogen-free saline to yield doses of 0.125, 0.25, and 0.5 mg/kg body weight. Mice were administered LLDT-8 (p.o. or i.p.), methotrexate (Shanghai Hualian Pharmaceutical Co., Ltd, Shanghai, China), or Leigongteng Duodai Pian (LDP, extracts from TWHF; Fudan Forward Pharmaceutical Co., Ltd, Shanghai, China) starting 1 day before the booster immunization and continuing once daily to evaluate its preventive effects. Alternatively, LLDT-8 treatment started from day 14 after booster immunization and continued once daily to evaluate its therapeutic effects where indicated.
Cell Preparation. Mice were sacrificed, and their axillary and inguinal lymph nodes (LN) or spleens were removed aseptically. A single cell suspension was prepared, and cell debris and clumps were removed. Erythrocytes were lysed with Tris-buffered ammonium chloride. Mononuclear cells were washed and resuspended in RPMI 1640 media supplemented with 10% fetal bovine serum, 100 U of penicillin/ml, and 100 μg of streptomycin/ml.
A B cell-depleted population was prepared by immunomagnetic negative selection as described by Yang et al. (2002). In brief, cells were incubated with magnetic particles bound to goat anti-mouse immunoglobulin (Ig) (QIAGEN, Valencia, CA), followed by removing cell-bound magnetic particles with a rare earth magnet (Polysciences, Inc., Warrington, PA).
Cell Proliferation. B cell-depleted LN cells (5 × 106/ml) were cultured in triplicate for 72 h with medium or 10 μg of CII/ml in 96-well flat bottom plates. Cells were pulsed with 0.25 μCi of [3H]-thymidine/well for 24 h and harvested onto glass fiber filters. The incorporated radioactivity was counted using a Beta Scintillation Counter (MicroBeta Trilux; PerkinElmer Life Sciences, Boston, MA).
Cytokine Production. B cell-depleted LN cells (5 × 106/ml) were cultured in triplicate for 48 h with medium or 100 μg of CII/ml in 96-well flat bottom plates. Culture supernatants were collected, and murine IFN-γ and IL-2 concentrations were determined by enzyme-linked immunosorbent assay (ELISA) (BD PharMingen, San Diego, CA).
Anticollagen Antibodies Detection. Sera were collected, and ELISA was used to detect antibodies to collagen. In brief, wells of flat-bottom plates were coated with 50 μg of CII/ml at 37°C for 1 h and then washed. Diluted samples were added for 1 h. Plates were washed and incubated for 30 min with goat anti-mouse Ig [IgG (H+L), IgG1, or IgG2a] coupled to horseradish peroxidase. After developing for 20 min with 0.1 mg of tetramethylbenzidine/ml and 0.1% H2O2 in 0.05 M citrate buffer (pH 5.4), the reactions were stopped by adding 1 M H2SO4. The optical density values at 450 and 570 nm were read using a microplate reader (Spectra MAX 190). As reference, a serum pool from DBA/1 mice exhibiting typical arthritis was used as standard and arbitrarily given a value of 300 U/ml.
Reverse Transcription-Polymerase Chain Reaction and Real-Time Polymerase Chain Reaction Analysis. B cell-depleted LN cells and spleen cells or joint homogenates were lysed using TRIzol reagent (Invitrogen, Carlsbad, CA). Total RNA was isolated, reverse transcribed, and polymerase chain reaction (PCR) was amplified using specific primers. Reverse transcription-PCR products were visualized by electrophoresis through 1% agarose gels containing ethidium bromide. Relative quantitation with real-time PCR was performed with SYBR Green PCR reagents (QIAGEN) and a continuous fluorescence detection system (MJ Research, Waltham, MA) according to the manufacturers' instructions. The mRNA levels were normalized to those of hypoxanthine-guanine phosphoribosyl-transferase.
NO Production and Measurement. Spleen cells (1 × 107/ml) were stimulated with 50 U of IFN-γ/ml (PharMingen) or 1 μg of lipopolysaccharide (LPS)/ml (Sigma) for 24 h in 96-well flat bottom plates. The production of NO was determined by assaying culture supernatant for NO2- as described previously (Zhou et al., 2006).
Determination of the Arginase Activity. Spleen cells were lysed, and the arginase activity was measured as described previously (Corraliza et al., 1994). One unit of enzyme activity is defined as the amount of enzyme that catalyzes the formation of 1 μmol of urea/min.
Flow-Cytometry Analysis of Mac-1+Cells. Spleen cells were stained with fluorescein isothiocyanate-conjugated anti-mouse CD11b monoclonal antibody (Mac-1+) (PharMingen) and analyzed on a FACSCalibur (BD Biosciences, San Jose, CA).
Histological Examination. Mice were sacrificed, and their hind limbs were removed and fixed in 10% formalin, then decalcified, and embedded in paraffin. Sections (5 μm) were stained with H&E. The histological damage was assessed and calculated using a 0 to 4 severity grade scoring system by two pathologists who were blinded to the experiments (0 = normal joint structure; 1 = mild changes, synovitis, and pannus front with few discrete cartilage focal erosions; 2 = moderate changes, accompanying loss of large areas of cartilage, eroding pannus front, and synovial hyperplasia with infiltrating mononuclear cells and polymorphonuclear cells; 3 = severe synovitis, cartilage and bone erosion; and 4 = total destruction of joint architecture).

Administration of LLDT-8 prevented the histological damage in CIA mice. CIA was induced, and mice were treated with 0.5 mg of LLDT-8/kg as described in Fig. 1. Hind joints of mice were subjected to histopathological examination. All the sections were stained with H&E. A, joint of normal mouse. B, joint of vehicle-treated CIA mouse; severe bone and cartilage surface erosion was observed. C, joint of CIA mouse treated with 0.5 mg of LLDT-8/kg (original magnification, ×4). D, histology score index was calculated. Data are mean ± S.D. (n = 8 mice/group). ***, p < 0.001 versus vehicle control.
Statistical Analysis. Data are presented as mean ± S.E.M. or mean ± S.D. where indicated. One-way analysis of variance followed by Dunnett's post-test or Student's t test was used to determine difference between two groups where appropriate. A p value <0.05 was considered significant.
Results
Administration of LLDT-8 Reduced the Incidence and Severity of CIA. Arthritis was elicited in DBA/1 mice by s.c. immunization twice with CII. Joint swelling was evident from day 4 after booster immunization, and almost 100% of mice developed arthritis when treated with vehicle alone. Figure 1, A and B, showed that administration of LLDT-8 significantly and dose-dependently reduced the incidence and severity of CIA, respectively. Pathologic damage occurs with the ongoing disease process. Severe cartilage and bone erosions accompanied by high numbers of lymphocyte infiltration were observed in vehicle-treated CIA mice (Fig. 2B). However, this joint damage was dramatically blocked in 0.5 mg/kg LLDT-8-treated mice (Fig. 2, C and D).
To assess the long-term effectiveness of LLDT-8, 3-month treatment experiments were performed. The p.o. administrations of LLDT-8 (0.125, 0.25, and 0.5 mg/kg, starting from 1 day before booster immunization) consistently attenuated the severity of CIA compared with vehicle-treated mice (Fig. 3A). LDP is a prescription drug for treating RA in China. The preventive effect of LLDT-8 at the dose of 0.25 mg/kg was similar to LDP at 20 mg/kg in CIA mice. We also tested the therapeutic effect of LLDT-8 after the establishment of the disease. CIA mice were treated with 0.5 mg of LLDT-8/kg starting from day 14 after booster immunization. The reduction of disease severity occurred from day 5 after LLDT-8 treatment, and the inhibitory profile was persistent during the 3-month observation (Fig. 3B).
Treatment with LLDT-8 Suppressed CII-Specific Immune Responses. To assess the effects of LLDT-8 on humoral immune response against CII, the concentrations of serum anti-CII antibodies were determined. The antibodies were barely detected in the sera of normal mice but were markedly produced in CIA mice. As shown in Fig. 4A, the induction of anti-CII IgG, IgG1, and IgG2a production was inhibited by LLDT-8. The level of anti-CII IgG2a was more significantly reduced than anti-CII IgG1 by LLDT-8 treatments (0.125, 0.25, and 0.5 mg/kg), displaying the 86, 78, and 93% reductions for IgG2a versus the 70, 77, and 85% reductions for IgG1 at the indicated doses, respectively.
To assess the effects of LLDT-8 on cell-mediated immune responses against CII, the recall proliferation of LN cells from CIA mice was examined. The results in Fig. 4B showed that LN cells from vehicle-treated mice well proliferated irrespective of CII stimulation, whereas the proliferation capacity of LN cells was greatly attenuated in LLDT-8-treated CIA mice.
Because T cells and Th1-type cytokines regulate the susceptibility and development of CIA (Myers et al., 1997), we investigated the capacity of LN cells to produce IFN-γ and IL-2 with CII stimulation. B cell-depleted LN cells produced large amounts of IFN-γ and IL-2 in CIA mice, but the increased production of both cytokines was reduced in LLDT-8-treated CIA mice (Fig. 4, C and D). Importantly, IFN-γ production seemed to be more susceptible to LLDT-8 treatment than IL-2 production. IL-4 was undetectable in our culture conditions (data not shown). Notably, after the disease was established, LLDT-8 treatment also suppressed IgG production (Fig. 4E), cell proliferation (Fig. 4F), and IFN-γ and IL-2 productions (Fig. 4, G and H).

Long-term effect of LLDT-8 treatment in CIA mice. A, prevention of the arthritis onset by LLDT-8 treatment. Mice were treated with LLDT-8 (p.o.) starting from 1 day before booster immunization. The severity of arthritis was evaluated, and representative data are shown; n = 12 mice/group. According to the statistical analysis at days 14, 42, and 90 after booster immunization, the severity in drug-treated mice was significantly reduced (p < 0.05) compared with vehicle-treated group. B, amelioration of the established arthritis by LLDT-8 treatment. Mice were treated with LLDT-8 (i.p.) starting from day 14 after booster immunization. The severity of arthritis was evaluated. Data are mean ± S.D. *, p < 0.05, **, p < 0.01, ***, p < 0.001 versus vehicle-treated control mice; n = 12 mice/group.
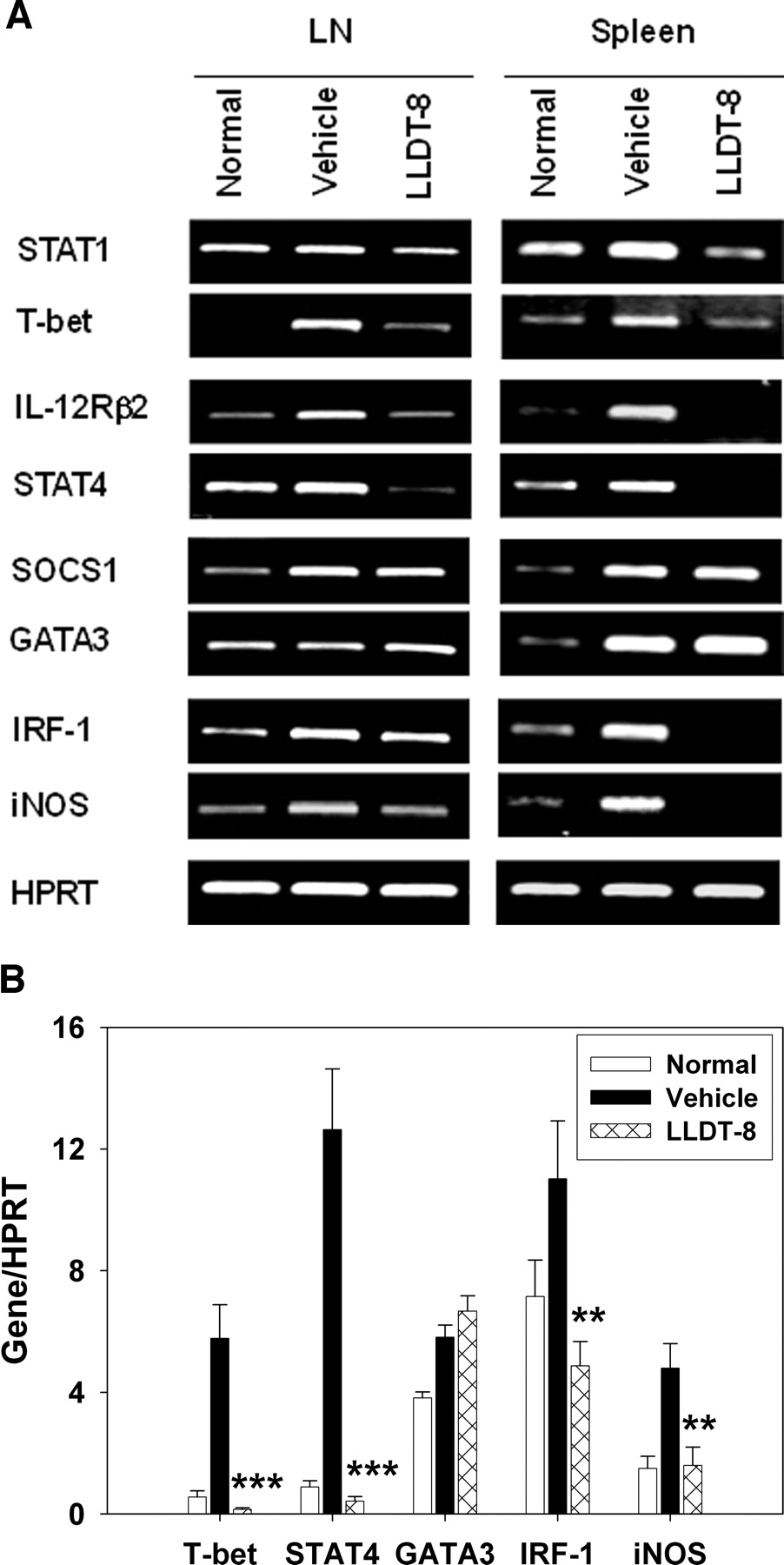
LLDT-8 Inhibited IFN-γ Production Pathway and iNOS Pathway in CIA Mice. To show the effect of LLDT-8 on IFN-γ signaling in CIA mice, PCR analysis was done in LN cells and spleen cells. Results are summarized in Fig. 5. Administration of LLDT-8 (0.5 mg/kg, i.p.) significantly reduced the transcripts of STAT1, T-bet, IL-12 receptor β2 (IL-12Rβ2), and STAT4 in IFN-γ production pathway. There was no significant difference for suppressor of cytokine signaling (SOCS) 1 and GATA binding factor 3 (GATA3) expressions between vehicle-treated and LLDT-8-treated mice (Fig. 5A). The T-bet, STAT4, and GATA3 expressions in LN cells were analyzed with real-time PCR. Similar inhibitory activity of LLDT-8 was observed (Fig. 5B).
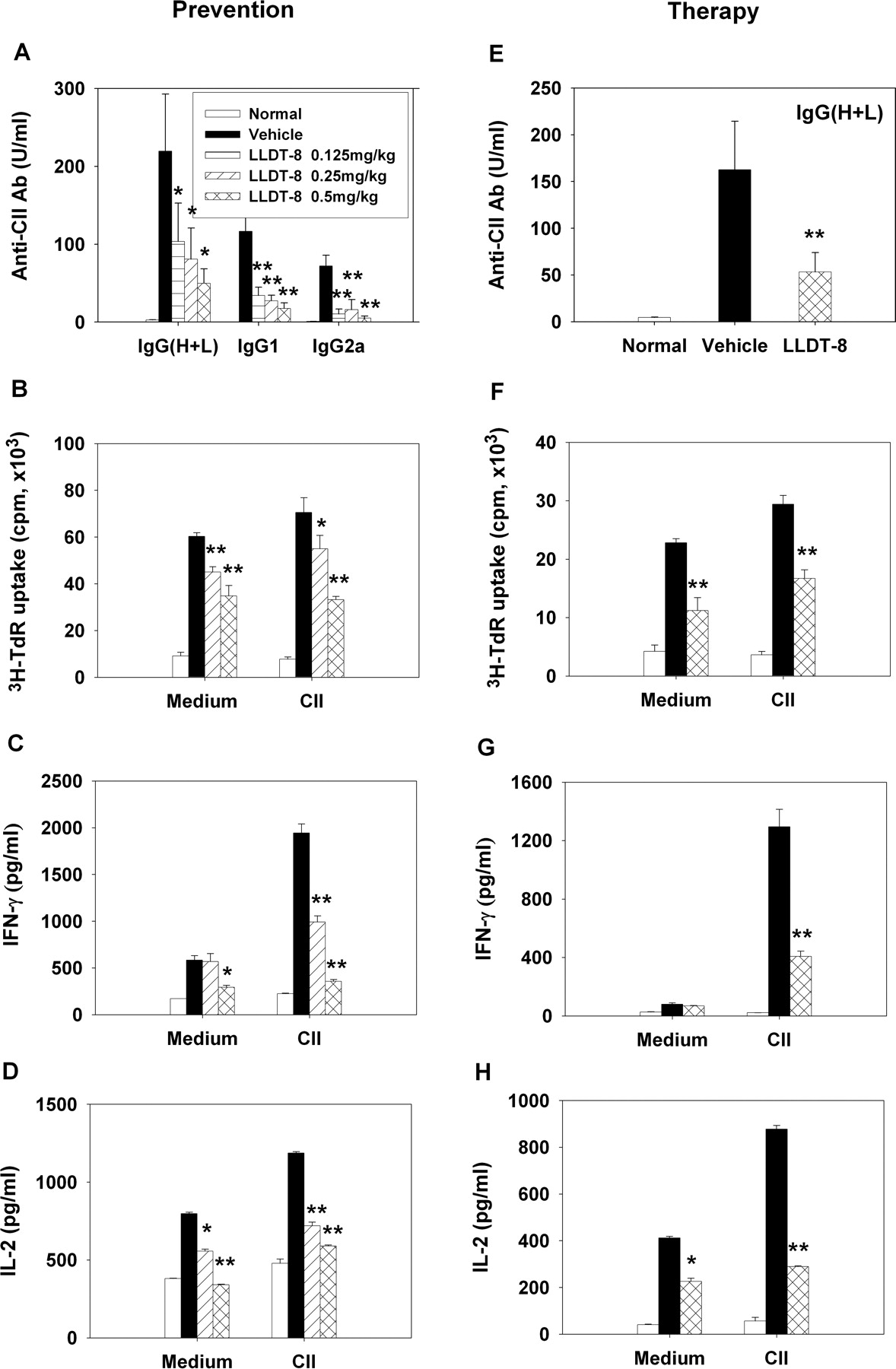
Inhibition of CII-specific immune responses by LLDT-8 treatment. Arthritis was induced, and mice were treated with LLDT-8 to prevent the onset of disease (A-D) or to attenuate the established disease (E-H). A and E, sera from mice were collected. The concentrations of anti-CII IgG, IgG1, and IgG2a were determined by ELISA, and data were calculated as arbitrary units; n = 12 mice/group. B and F, B cell-depleted LN cells were cultured in triplicate with medium or 10 μg of CII/ml for 72 h and pulsed with [3H]thymidine. The incorporated radioactivity was measured; n = 6 mice/group. C, D, G, and H, B cell-depleted LN cells were cultured in triplicate with medium or 100 μg of CII/ml for 48 h. Supernatants were assayed for IFN-γ (C and G) and IL-2 (D and H) by ELISA; n = 6 mice/group. Data are mean ± S.E.M. of two independent experiments. *, p < 0.05, **, p < 0.01 versus vehicle-treated mice. The treatment period of LLDT-8 was 2 weeks in A through D and 4 weeks in E through H.
In our previous study, LLDT-8 displayed a strong inhibition on NO production in both IFN-γ signaling pathway and LPS signaling pathway (Zhou et al., 2006). Here, LLDT-8 greatly reduced the IRF-1 and iNOS mRNA transcriptions in IFN-γ/STAT1/IRF-1/iNOS pathway in CIA mice (Fig. 5, A and B). Furthermore, the unfractionated spleen cells were stimulated ex vivo with IFN-γ (Fig. 6, A and E) or LPS (Fig. 6, B and F) to induce NO production. NO generation remarkably increased in vehicle-treated CIA mice over the basal level in normal mice. Treatment with 0.5 mg of LLDT-8/kg effectively blocked this increase.
Arginase competes with iNOS for arginine. The activity of arginase was determined in spleen cells from CIA mice. As Fig. 6C shows, the enzyme activity increased in vehicle-treated CIA mice. Administration of LLDT-8 (0.5 mg/kg, i.p.) potently blocked this abnormal increase and retained the activity of arginase at the basal level as that in the normal control mice. After the onset of arthritis, mice were treated with LLDT-8, and the arginase activity was blocked as well (Fig. 6G). LLDT-8 treatment did not affect the number of macrophages compared with that in vehicle-treated mice (Fig. 6, D and H).
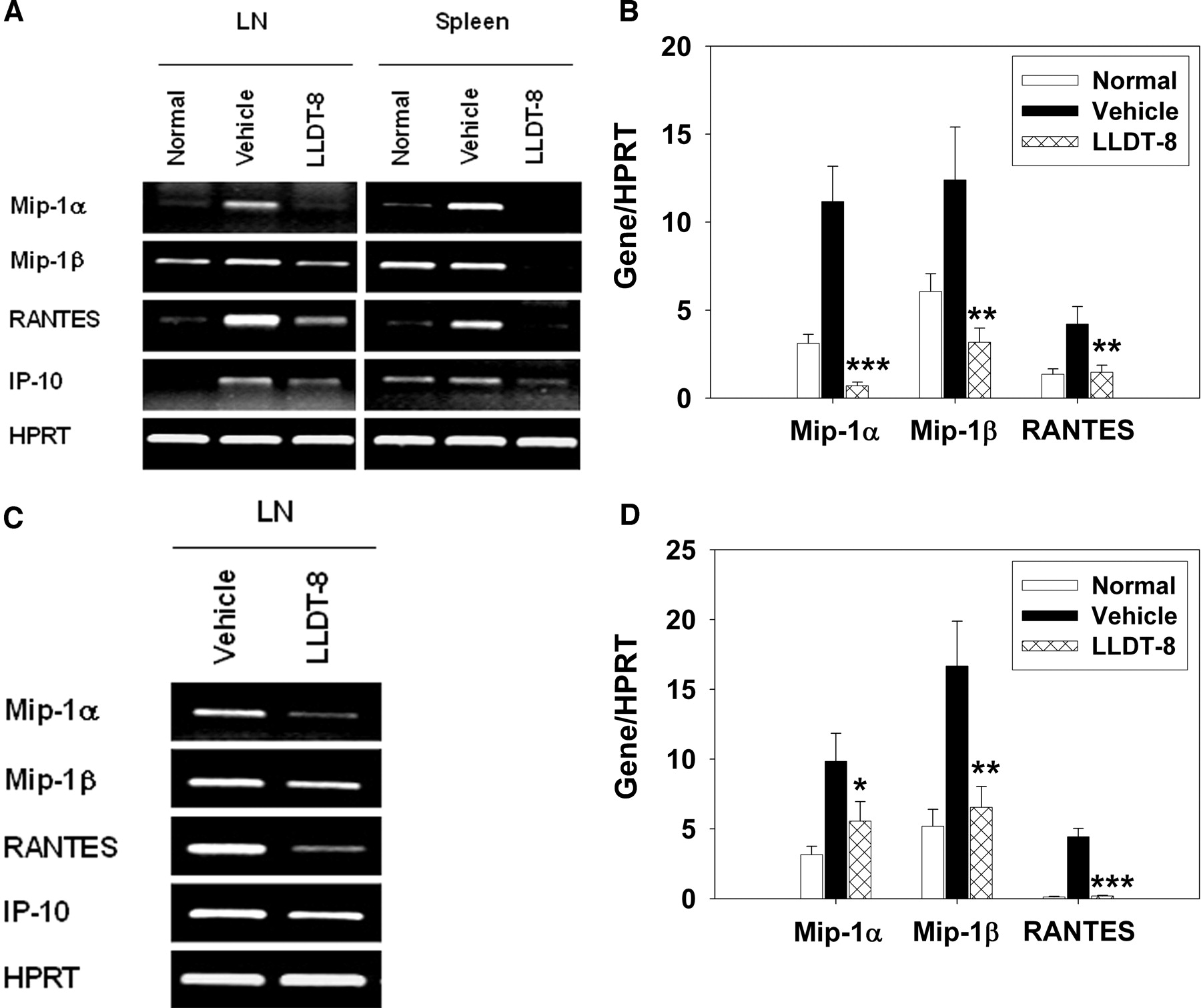
LLDT-8 Blocked IFN-γ-Induced Chemokine Expressions and Inhibited Their Receptor Expressions. RA is an inflammatory disorder characterized by infiltration of leukocytes into the synovial tissue and synovial fluid of the joints. IFN-γ can regulate the leukocyte trafficking through induction of various chemokine genes, including Mip-1α, Mip-1β, RANTES, and IP-10 (Schroder et al., 2004). To understand the mechanisms underlying the effects of LLDT-8 in leukocyte infiltration, expressions of a variety of chemokines were compared by PCR analysis in LN cells and/or spleen cells from CIA mice. As shown in Fig. 7, the transcripts of Mip-1α, Mip-1β, RANTES, and IP-10 were less in naive DBA/1 mice but dramatically increased in CIA mice. Administration of LLDT-8 (0.5 mg/kg) significantly reduced the mRNA expression of these chemokines (Fig. 7, A and C). The suppression of chemokine expressions by LLDT-8 was further shown with real-time PCR analysis in LN cells (Fig. 7, B and D).
Elevated expression of CCR5 and CXCR3 has been reported on synovial fluid T cells in juvenile RA, associated with a high IFN-γ level (Wedderburn et al., 2000). The expressions of chemokine receptors were analyzed with PCR in LN, spleen, or joint tissues. Up-regulated mRNA expressions of CCR1, CCR5, and CXCR3 were observed in CIA mice over that in naive mice, which is in line with the presence of their ligands. Treatment with LLDT-8 (0.5 mg/kg) decreased their expressions to basal levels before (Fig. 8, A and B) or after (Fig. 8, C and D) the disease was established.
Discussion
Here we first report the use of LLDT-8, a novel analog of triptolide, as an antiarthritic therapeutic agent in CIA mice. LLDT-8 reduced the disease incidence, severity, and CII-specific immune responses via blocking IFN-γ signaling pathways, including IFN-γ/STAT1/T-bet/IFN-γ pathway, IL-12/IL-12Rβ2/STAT4/IFN-γ pathway, IFN-γ/STAT1/IRF-1/iNOS pathway, and IFN-γ-regulated chemokine pathway. The expressions of chemokine receptors were suppressed as well.
CIA is an animal model for RA. In the present study, we found that LLDT-8 significantly improved the clinical and histological manifestations of CIA. LLDT-8 could prevent the onset and continued presence of arthritis, and LLDT-8 showed beneficial effects even after the arthritis was established. Only approximately 10% of mice developed mild arthritis after cessation of LLDT-8 treatment (R. Zhou, unpublished observations). Together, our data suggested the therapeutic potential of LLDT-8 for the clinical treatment of RA patients. Now, LLDT-8 is systematically evaluated as a candidate drug for RA in the preclinical study.
The onset of CIA often accompanies high levels of circulating autoantibody to initiate joint inflammation, especially of the IgG2a subclass (Myers et al., 1997). LLDT-8 reduced the IgG2a level, as well as total IgG and IgG1, displaying a stronger inhibition on IgG2a than on IgG1. Because IFN-γ mediates the switch of antibody production to IgG2a, it was possible that LLDT-8 reduced the IgG2a level through inhibiting the IFN-γ production. IgG2a is associated with Th1 responses, whereas IgG1 is associated with Th2 responses. Thus, LLDT-8 might attenuate both Th1 and Th2 immune responses against CII, leading to the suppression of humoral immune responses in CIA. LLDT-8 also inhibited the cellular immune responses against CII in CIA. The lymphocyte proliferation was impaired in LLDT-8-treated CIA mice, with no distinct difference in response to medium alone or CII. The “ineffectiveness” of CII stimulation might be attributed to that residual CII contained in antigen presenting cells in LN sufficient to activate T cell proliferation.
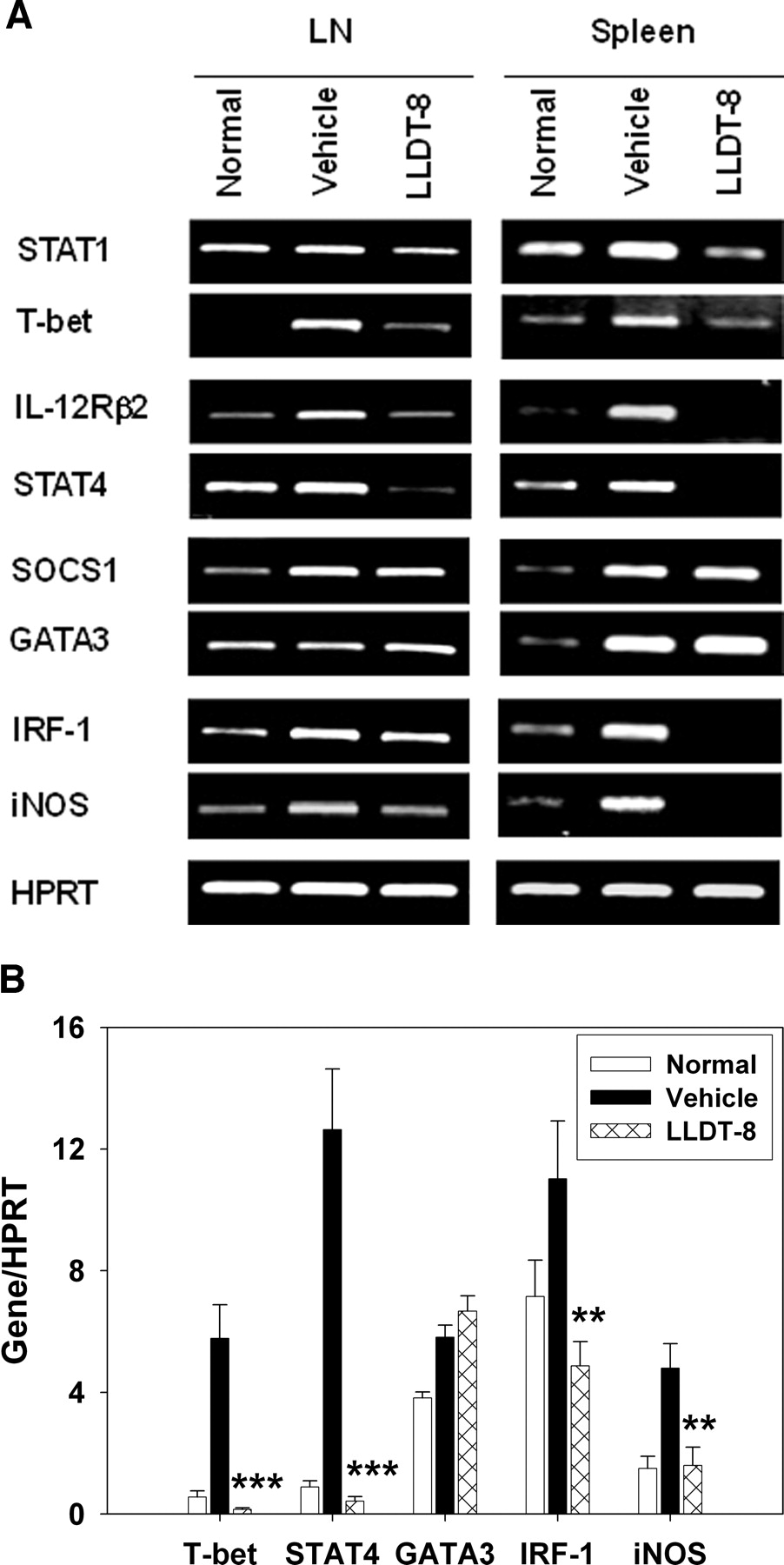
Effects of LLDT-8 on IFN-γ production pathway and iNOS pathway in CIA mice. To analyze the preventive effect of LLDT-8 on gene expression, B cell-depleted LN cells and spleen cells were prepared. A, total RNA was isolated and analyzed for mRNA expression with reverse transcription-PCR. Three independent experiments were performed that gave similar results. B, mRNA expressions in LN cells were analyzed with real-time PCR. Data are mean ± S.E.M. of two independent experiments. **, p < 0.01, ***, p < 0.001 versus vehicle-treated mice.
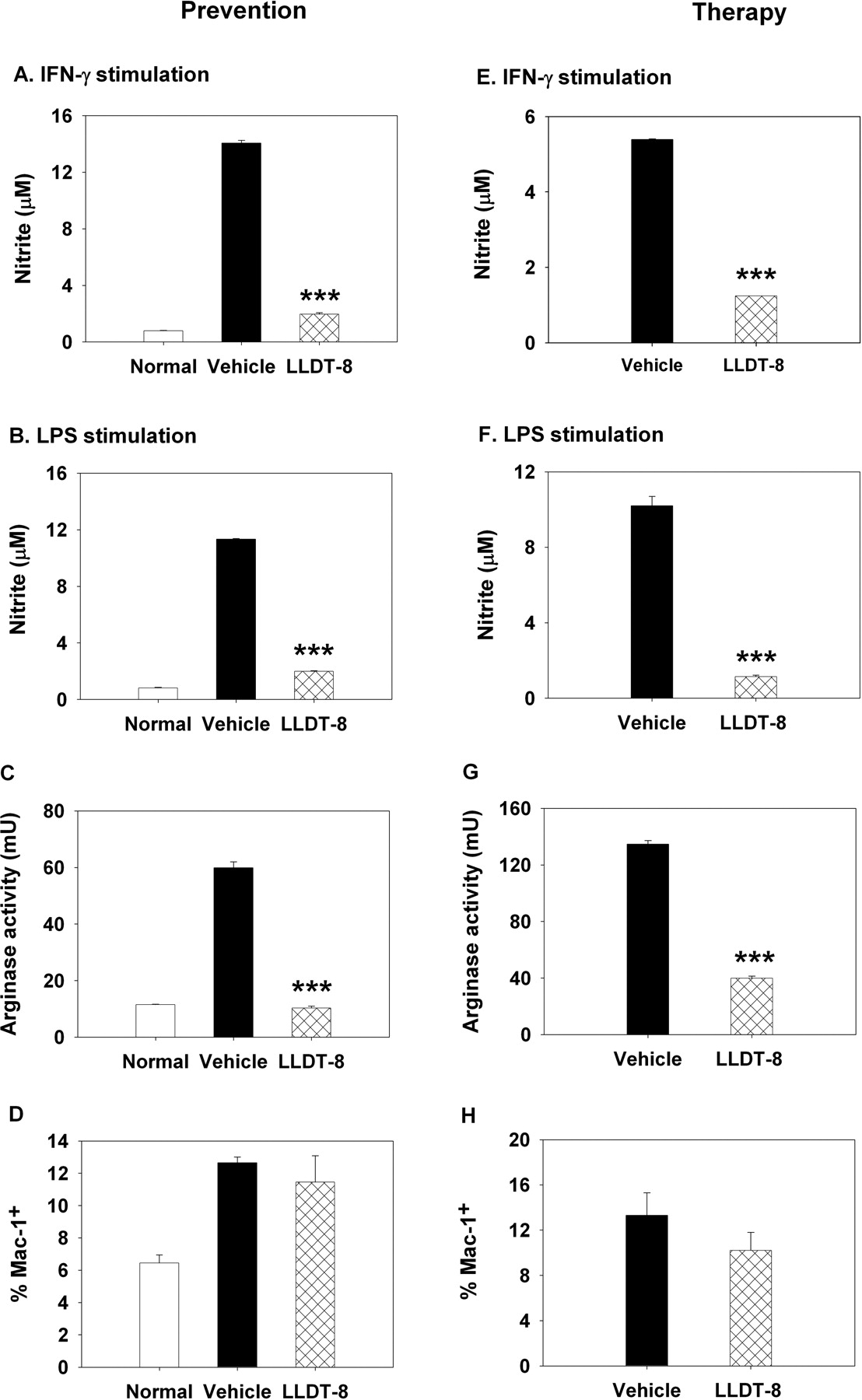
Effects of LLDT-8 on NO production and arginase activity. Arthritis was induced, and mice were treated with LLDT-8 to prevent the onset of disease (A-D) or to attenuate the established disease (E-H). Spleen cells were stimulated with 50 U of IFN-γ/ml (A and E) or 1 μg of LPS/ml (B and F) for 24 h. The supernatants were assayed for nitrite. Data are mean ± S.D. of triplicate cultures. C and G, spleen cells were lysed and assayed for arginase activity. Data are mean ± S.D. of triplicate samples. Three independent experiments are performed that give similar results. *, p < 0.05, **, p < 0.01, ***, p < 0.001 versus vehicle-treated mice. D and H, spleen cells were analyzed for Mac-1 by flow cytometry. Data are mean ± S.E.M. of three independent experiments.
IFN-γ plays a dual role in CIA, functioning as both a disease-limiting (Nakajima et al., 1990; Manoury-Schwartz et al., 1997; Vermeire et al., 1997) and a disease-promoting factor (Mauritz et al., 1988; Skurkovich and Skurkovich, 2003). Until now, no satisfactory explanation for the controversial effects of IFN-γ has been offered. Elimination of IFN-γ either by neutralizing antibodies or in IFN-γ-/- and IFN-γ receptor-/- mice leads to disease exacerbation (Manoury-Schwartz et al., 1997; Vermeire et al., 1997). IFN-γ may protect CIA through restraint from mycobacteria (in CFA)-stimulated expansion of the Mac-1+ cell population (Matthys et al., 1999), up-regulation of CD4+CD25+ T regulatory activity (Kelchtermans et al., 2005), or early prevention of the disease followed by late-stage disease exacerbation (Boissier et al., 1995). Meanwhile, it has been shown that IFN-γ may exacerbate CIA through many mechanisms, such as the production of inflammatory cytokines, NO, and antibodies against CII (Kageyama et al., 1998). In LLDT-8-treated CIA mice, the reduction in disease was principally correlated with decreased levels of IFN-γ, suggesting a critical role of IFN-γ in the LLDT-8 action. The IFN-γ/STAT1/T-bet/IFN-γ and IL-12/IL-12Rβ2/STAT4/IFN-γ pathways mediate the IFN-γ production (Grogan and Locksley, 2002). LLDT-8 reduced the mRNA expressions of STAT1, T-bet, IL-12Rβ2, and STAT4, providing the molecular basis for its inhibition of IFN-γ and the pathogenesis in CIA. Moreover, in the synovium of RA and experimental arthritis, macrophages express elevated levels of STAT1 and STAT4, followed by abnormal activation. The continuous STAT1 activation may be of pathogenic significance in the progression and persistence of RA (Frucht et al., 2000; Yokota et al., 2001). LLDT-8 reduced STAT1 expression in CIA mice, probably contributing to the suppression of arthritis. IL-12 regulates the magnitude of IFN-γ expression through a STAT4-dependent pathway, and arthritis is suppressed in STAT4-/- mice (Finnegan et al., 2002). LLDT-8 intensively blocked the transcription of IL-12Rβ2 and its downstream STAT4, which might lead to the suppression of arthritis. IFN-γ signaling is dependent on activated STAT1, and tyrosine phosphorylation of STAT1 is under feedback inhibition by SOCS1 (Naka et al., 1999). GATA3 positively regulates IL-4 production and negatively regulates Th1 responses (Zheng and Flavell, 1997). LLDT-8 showed no effect on SOCS1 and GATA3 transcriptions. Together, LLDT-8 significantly inhibited the IFN-γ production signaling in the Th1 response without enhancing Th2-type response.

Suppression of chemokine expressions by LLDT-8. Arthritis was induced, and mice were treated with LLDT-8 to prevent the onset of disease (A and B) or to attenuate the established disease (C and D). B cell-depleted LN cells and spleen cells were prepared. A and C, total RNA was isolated and analyzed with reverse transcription-PCR for mRNA expressions. Three independent experiments are performed that give similar results. B and D, mRNA expressions in LN cells were analyzed with real-time PCR. Data are mean ± S.E.M. of two independent experiments. *, p < 0.05, **, p < 0.01, ***, p < 0.001 versus vehicle-treated mice.
IFN-γ exacerbates CIA through a variety of mechanisms. iNOS locates at the downstream of IFN-γ/STAT1/IRF-1 signaling. The incidence and severity of CIA are significantly decreased in IRF-1-/- mice compared with IRF-l+/- mice (Tada et al., 1997). Nonselective inhibition of NOS suppresses the experimental arthritis (Stefanovic-Racic et al., 1994). LLDT-8 inhibited the IFN-γ/STAT1/IRF-1/iNOS pathway, contributing to the suppression of arthritis. Excessive production of NO generated by iNOS has been described in patients with RA (Farrell et al., 1992), and its concentration was related to disease activity. We have shown that LLDT-8 reduces NO production and iNOS expression by inhibiting IFN-γ-triggered IRF-1 expression and LPS-triggered mitogen-activated protein kinase phosphorylation and nuclear factor κB activation in Raw 264.7 cells (Zhou et al., 2006). In this study, the NO production was greatly attenuated in LLDT-8-treated mice, irrespective of the stimuli difference (IFN-γ or LPS). The numbers of Mac-1+, the NO-producing cells in spleen, were almost the same in vehicle-treated and LLDT-8-treated CIA mice. Thus, the possibility of reducing Mac-1+ cell numbers to decrease NO production was excluded. Notably, when stimulated with the same amount of IFN-γ, the NO production was still significantly suppressed in LLDT-8-treated CIA mice. This further verified that the IFN-γ signaling pathway was impaired by LLDT-8 treatment. The expression of arginase II is up-regulated in synovial fluid cells from RA patients, participating in cell proliferation and tissue regeneration (Corraliza and Moncada, 2002). This might accelerate joint distortion. LLDT-8 retarded the abnormal increase of arginase activity and prevented the onset of arthritis. Unexpectedly, there was no significant increase for Mac-1+ cell number, NO production, and arginase activity in draining LN cells (data not shown).

Suppression of chemokine receptor expressions by LLDT-8. Arthritis was induced, and mice were treated with LLDT-8 to prevent the onset of disease (A and B) or to attenuate the established disease (C and D). B cell-depleted LN cells and spleen cells or homogenates from joint tissues were prepared. A and C, total RNA was isolated and analyzed with reverse transcription-PCR for mRNA expressions. Three independent experiments are performed that give similar results. B and D, mRNA expressions in LN cells were analyzed with real-time PCR. Data are mean ± S.E.M. of two independent experiments. *, p < 0.05, **, p < 0.01, ***, p < 0.001 versus vehicle-treated mice.
In the pathogenesis of RA, leukocytes infiltrate into the synovial tissue. IFN-γ induces Mip-1α, Mip-1β, and IP-10 in the presence of STAT1 (Gil et al., 2001). IRF-1 response elements have been identified in the promoters of IP-10 (Nazar et al., 1997). Thus, IFN-γ may induce these chemokine expressions also via the IFN-γ/STAT1/IRF-1 pathway. Mip-1α and RANTES are important for the leukocyte influx into RA joints. The expressions of Mip-1α, Mip-1β, RANTES, and IP-10 were suppressed by LLDT-8 in CIA mice. These results suggested that LLDT-8 reduced attraction of leukocyte infiltration into inflammatory sites. CCR5 has been detected in the RA synovium. CXCR3 is expressed in both allergic and synovial infiltrates (Qin et al., 1998). Up-regulation and activation of CCR1 and CCR5 play a role in macrophage and endothelial cell infiltration in rat adjuvant-induced arthritis (Shahrara et al., 2003). In the LLDT-8-treated CIA mice, the CCR1, CCR5, and CXCR3 mRNA expressions were reduced, especially in joint tissue. Thus, LLDT-8 might block lymphocyte infiltration into joints.
Although LLDT-8 was shown to be effective for the treatment of CIA, something still remained unclear. Because T-bet and IRF-1 act downstream of STAT1, and STAT4 acts downstream of IL-12Rβ2, additional studies are required to clarify whether LLDT-8 suppresses IFN-γ signaling at multiple points or just at the initial steps (STAT1 and IL-12Rβ2). Activation of T cells via major histocompatability complex-peptide-T cell receptor can induce IFN-γ production, but this signaling cascade is not very clear. Although LLDT-8 exhibited weak effect on antigen presentation (R. Zhou, unpublished observations), we could not exclude the T cell receptor pathway from the effect of LLDT-8 on IFN-γ signaling.
In conclusion, our results suggested that LLDT-8 had an antiarthritic effect in CIA mice, and the beneficial effects were tightly associated with blockade of IFN-γ signaling pathways. LLDT-8 may have therapeutic potentials in the treatment of RA and other autoimmune diseases.
Acknowledgments
We thank the research group of Dr. Jin Ren (Shanghai Institute of Materia Medica) for providing valuable technical assistance with histology.
Footnotes
-
This work was supported by a grant from the Knowledge Innovation Program of Chinese Academy of Sciences (KSCX2-SW-202).
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.101113.
-
ABBREVIATIONS: TWHF, T. wilfordii Hook F; LLDT-8, (5R)-5-hydroxytriptolide; CIA, collagen-induced arthritis; RA, rheumatoid arthritis; CII, type II bovine collagen; Th, T helper; IFN-γ, interferon-γ; IL, interleukin; STAT, signal transducer and activator of transcription; T-bet, T-box transcription factor; IRF, interferon regulatory factor; iNOS, inducible nitric-oxide synthase; NO, nitric oxide; Mip, macrophage inflammatory protein; RANTES, regulated on activation normally T-cell expressed and secreted; IP-10, inducible protein 10; CCR, C-C chemokine receptor; CXCR, C-X-C chemokine receptor; CFA, Freund's complete adjuvant; LDP, Leigongteng Duodai Pian is the prescription drug of the extracts from TWHF in China; LN, lymph node(s); Ig, immunoglobulin; ELISA, enzyme-linked immunosorbent assay; PCR, polymerase chain reaction; LPS, lipopolysaccharide; IL-12Rβ2, IL-12 receptor β2; SOCS, suppressor of cytokine signaling; GATA3, GATA binding factor 3.
- Received January 9, 2006.
- Accepted March 27, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}